

Behaviorally consequential astrocytic regulation of neural circuits
- PMID: 33385325
- PMCID: PMC7897322
- DOI: 10.1016/j.neuron.2020.12.008
Behaviorally consequential astrocytic regulation of neural circuits
Abstract
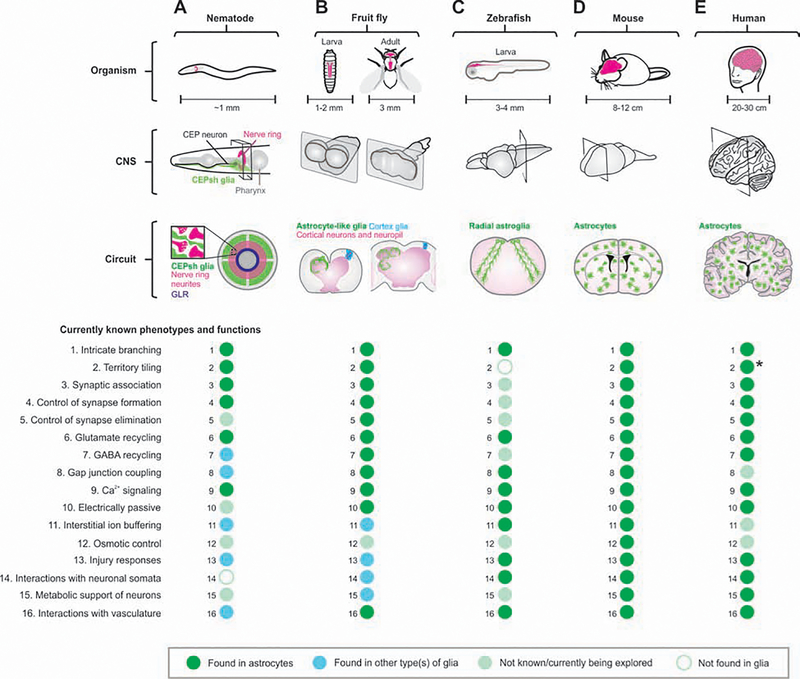
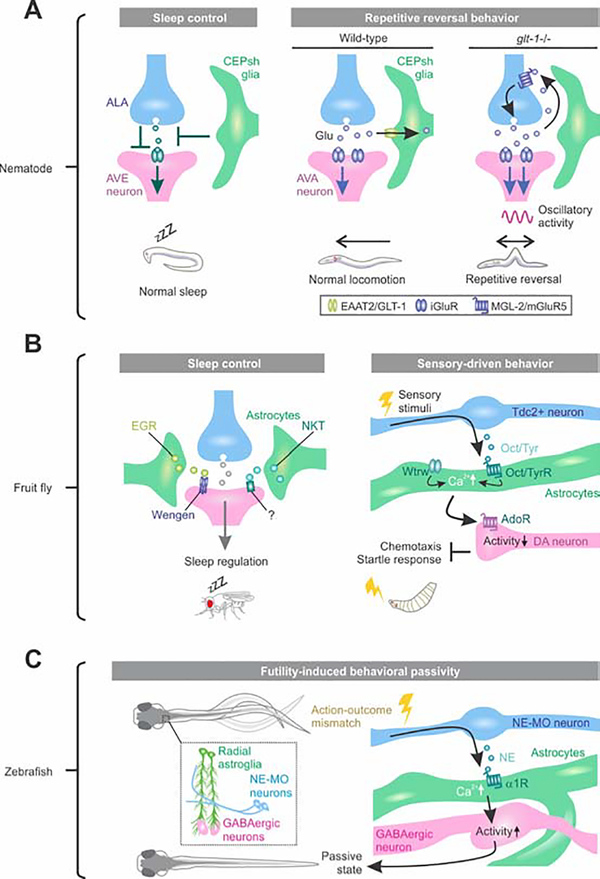
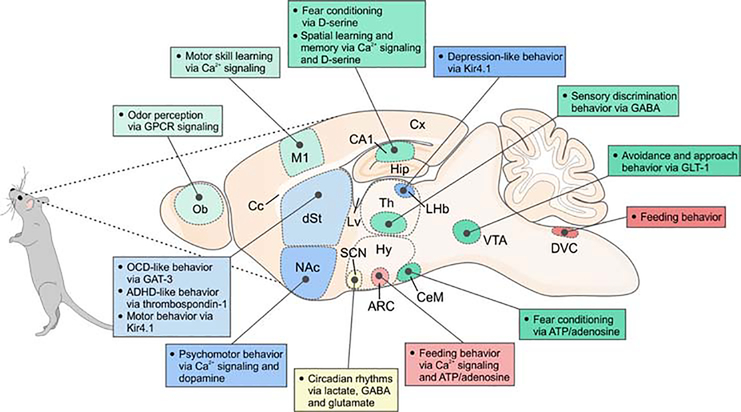
Astrocytes are a large and diverse population of morphologically complex cells that exist throughout nervous systems of multiple species. Progress over the last two decades has shown that astrocytes mediate developmental, physiological, and pathological processes. However, a long-standing open question is how astrocytes regulate neural circuits in ways that are behaviorally consequential. In this regard, we summarize recent studies using Caenorhabditis elegans, Drosophila melanogaster, Danio rerio, and Mus musculus. The data reveal diverse astrocyte mechanisms operating in seconds or much longer timescales within neural circuits and shaping multiple behavioral outputs. We also refer to human diseases that have a known primary astrocytic basis. We suggest that including astrocytes in mechanistic, theoretical, and computational studies of neural circuits provides new perspectives to understand behavior, its regulation, and its disease-related manifestations.
Keywords: Caenorhabditis elegans; Danio rerio; Drosophila melanogaster; Mus musculus; astrocyte; behavior; genetic disorders; glia; microcircuit; neuronal circuit.
Copyright © 2020 Elsevier Inc. All rights reserved.
Figures
References
-
- Adamsky A, Kol A, Kreisel T, Doron A, Ozeri-Engelhard N, Melcer T, Refaeli R, Horn H, Regev L, Groysman M, et al. (2018). Astrocytic Activation Generates De Novo Neuronal Potentiation and Memory Enhancement. Cell 174, 59–71. - PubMed
-
- Agulhon C, Fiacco TA, and McCarthy KD (2010). Hippocampal short- and long-term plasticity are not modulated by astrocyte Ca2+ signaling. Science 327, 1250–1254. - PubMed
Publication types
MeSH terms
Grants and funding
- MC_PC_17230/MRC_/Medical Research Council/United Kingdom
- R37 NS053538/NS/NINDS NIH HHS/United States
- R01 DA047444/DA/NIDA NIH HHS/United States
- MC_U105170643/MRC_/Medical Research Council/United Kingdom
- R01 NS053538/NS/NINDS NIH HHS/United States
- R35 NS111583/NS/NINDS NIH HHS/United States
- R01 AG061838/AG/NIA NIH HHS/United States
- WT_/Wellcome Trust/United Kingdom
- R01 NS037585/NS/NINDS NIH HHS/United States
- R21 NS115437/NS/NINDS NIH HHS/United States
- R01 MH127163/MH/NIMH NIH HHS/United States
- R01 NS107315/NS/NINDS NIH HHS/United States
- MC_PC_12009/MRC_/Medical Research Council/United Kingdom
- R35 NS105094/NS/NINDS NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
