

TET-dependent GDF7 hypomethylation impairs aqueous humor outflow and serves as a potential therapeutic target in glaucoma
- PMID: 33388417
- PMCID: PMC8058441
- DOI: 10.1016/j.ymthe.2020.12.030
TET-dependent GDF7 hypomethylation impairs aqueous humor outflow and serves as a potential therapeutic target in glaucoma
Abstract
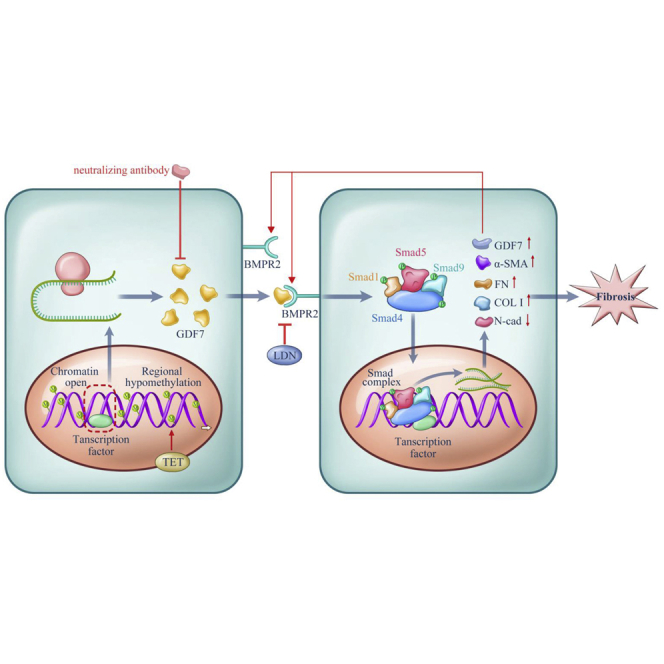
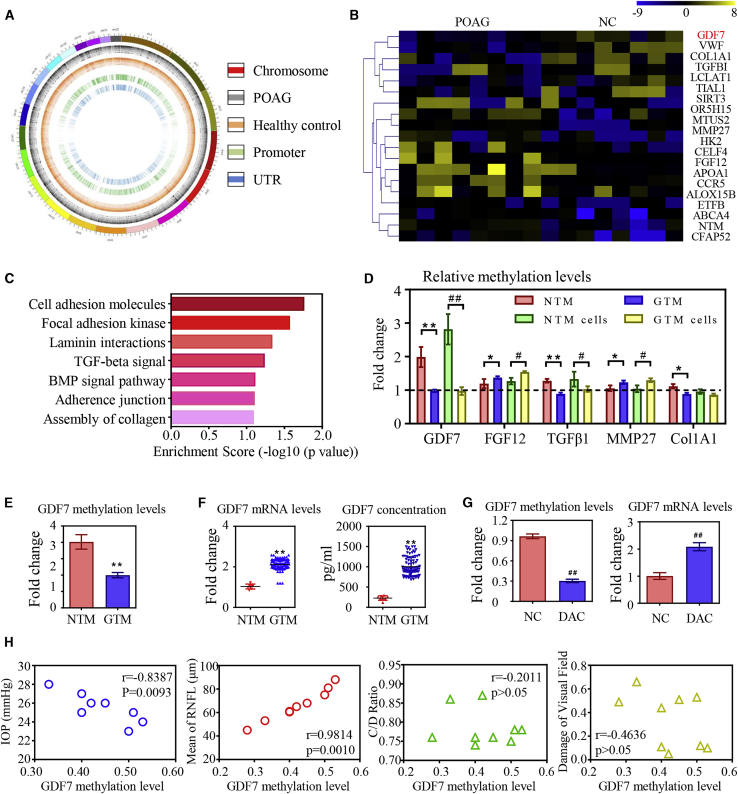
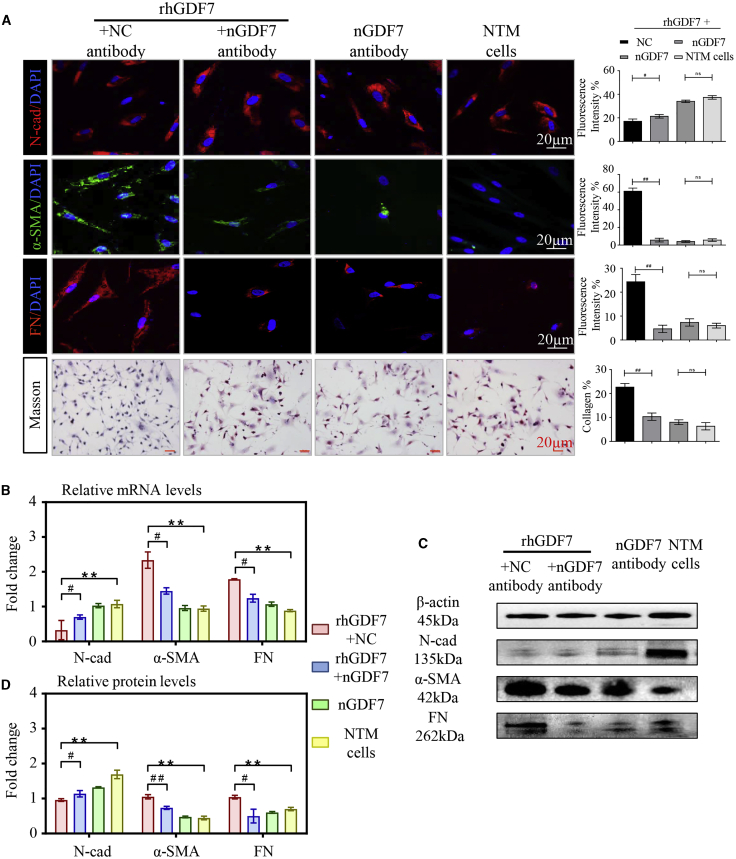
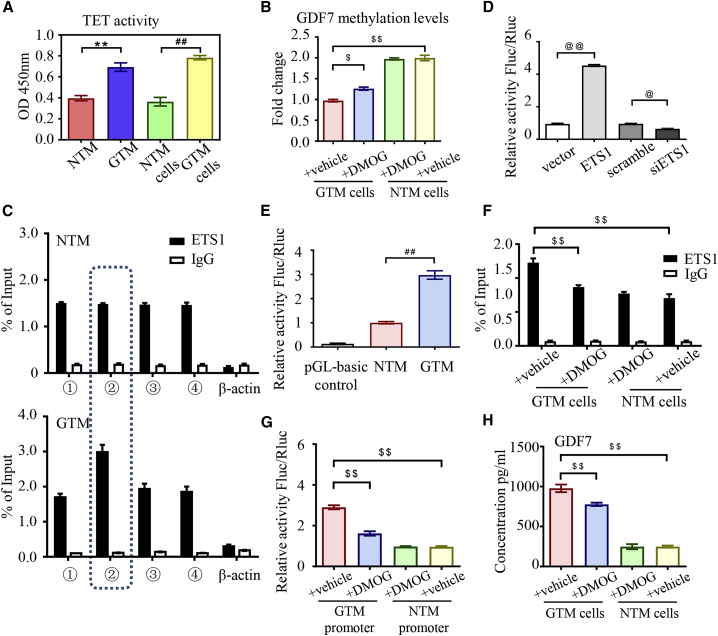
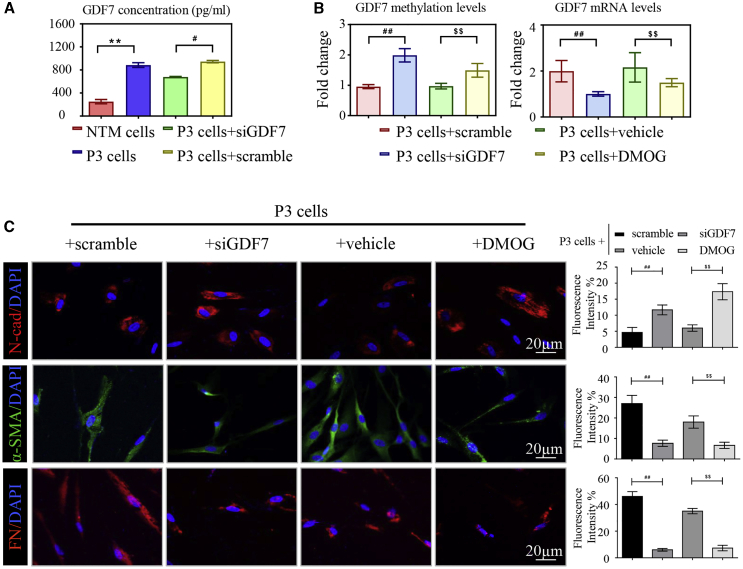
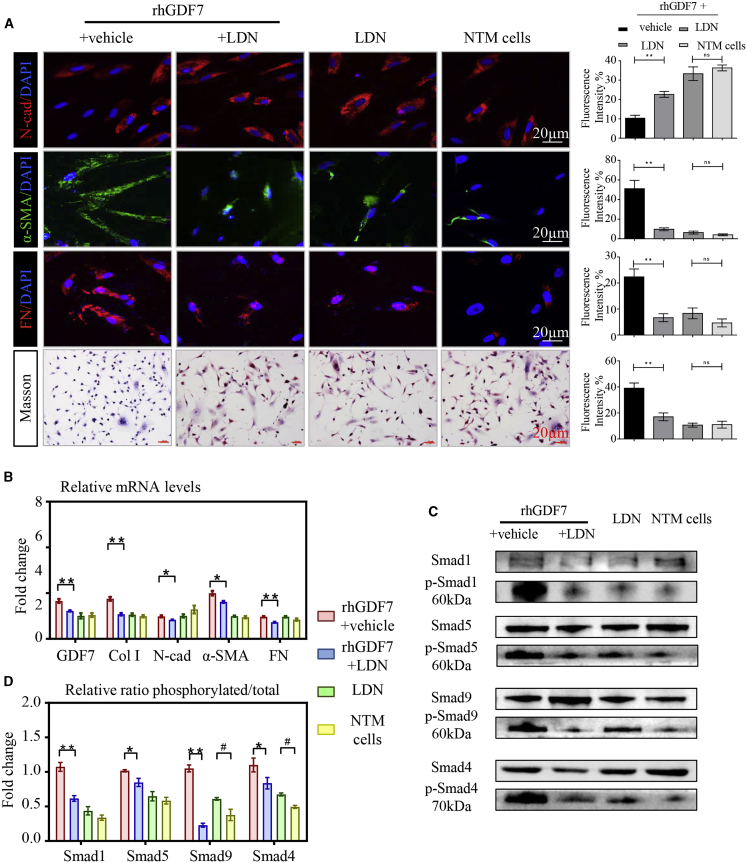
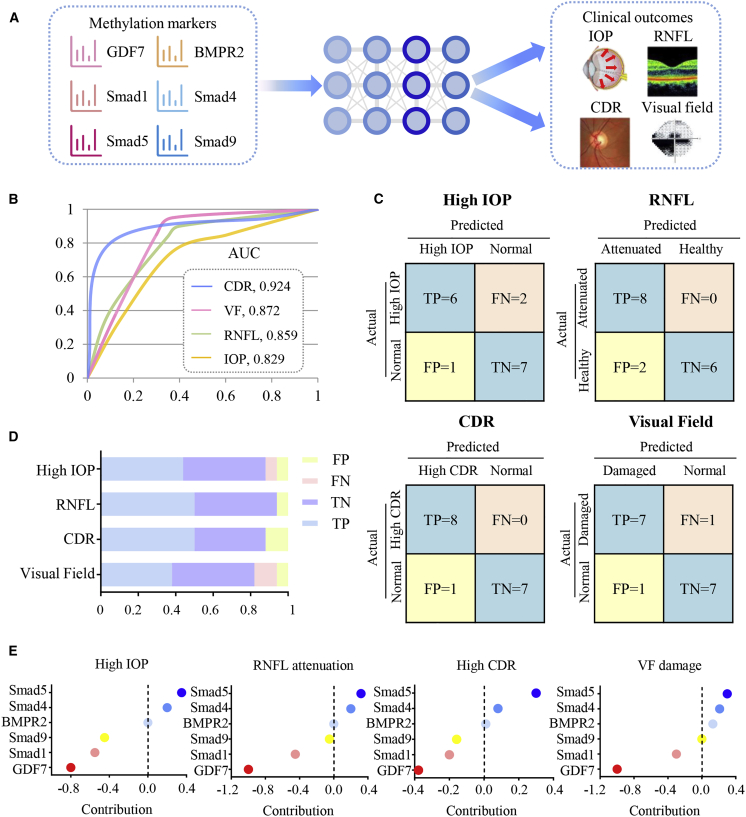
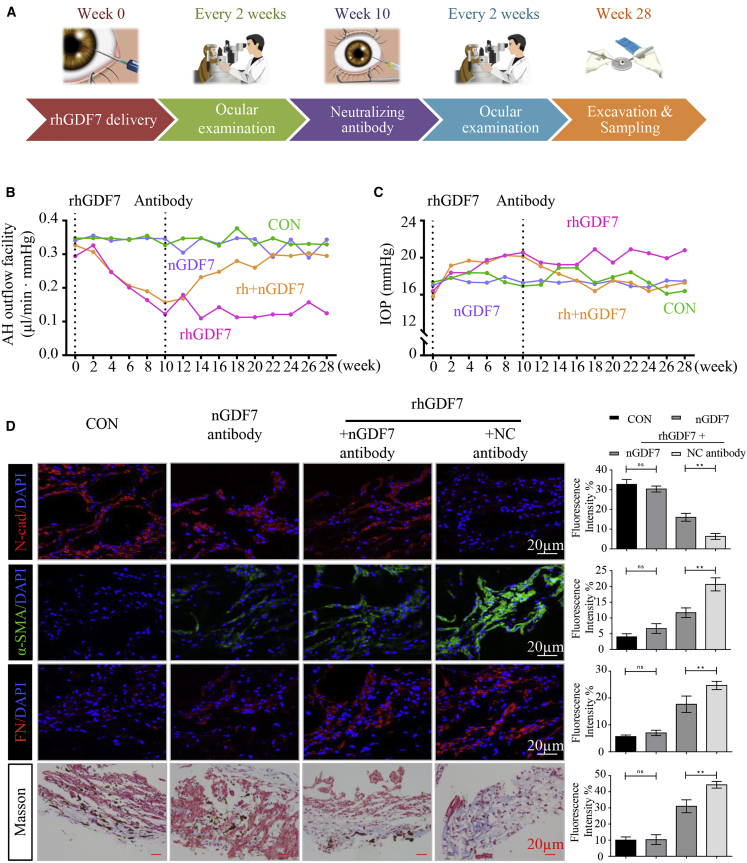
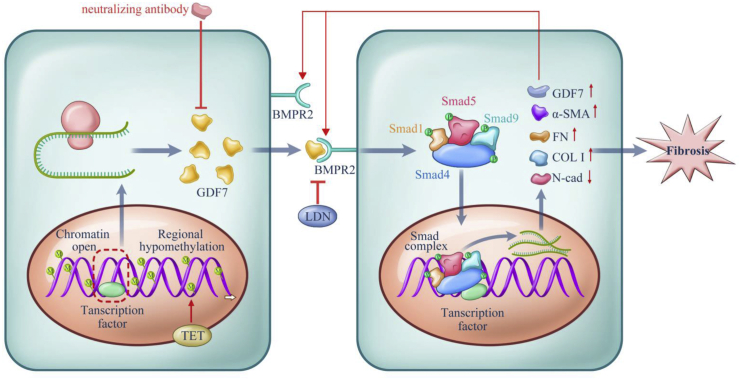
Glaucoma is the leading cause of irreversible vision loss, affecting more than 70 million individuals worldwide. Circulatory disturbances of aqueous humor (AH) have long been central pathological contributors to glaucomatous lesions. Thus, targeting the AH outflow is a promising approach to treat glaucoma. However, the epigenetic mechanisms initiating AH outflow disorders and the targeted treatments remain to be developed. Studying glaucoma patients, we identified GDF7 (growth differentiation factor 7) hypomethylation as a crucial event in the onset of AH outflow disorders. Regarding the underlying mechanism, the hypomethylated GDF7 promoter was responsible for the increased GDF7 production and secretion in primary open-angle glaucoma (POAG). Excessive GDF7 protein promoted trabecular meshwork (TM) fibrosis through bone morphogenetic protein receptor type 2 (BMPR2)/Smad signaling and upregulated pro-fibrotic genes, α-smooth muscle actin (α-SMA) and fibronectin (FN). GDF7 protein expression formed a positive feedback loop in glaucomatous TM (GTM). This positive feedback loop was dependent on the activated TET (ten-eleven translocation) enzyme, which kept the GDF7 promoter region hypomethylated. The phenotypic transition in TM fortified the AH outflow resistance, thus elevating the intraocular pressure (IOP) and attenuating the nerve fiber layer. This methylation-dependent mechanism is also confirmed by a machine-learning model in silico with a specificity of 84.38% and a sensitivity of 89.38%. In rhesus monkeys, we developed GDF7 neutralization therapy to inhibit TM fibrosis and consequent AH outflow resistance that contributes to glaucoma. The neutralization therapy achieved high-efficiency control of the IOP (from 21.3 ± 0.3 to 17.6 ± 0.2 mmHg), a three-fold improvement in the outflow facility (from 0.1 to 0.3 μL/min · mmHg), and protection of nerve fibers. This study provides new insights into the epigenetic mechanism of glaucoma and proposes an innovative GDF7 neutralization therapy as a promising intervention.
Keywords: DNA methylation; computational modeling; fibrosis; glaucoma; neutralizing antibody; trabecular meshwork.
Copyright © 2020 The Author(s). Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures
References
-
- Tham Y.C., Li X., Wong T.Y., Quigley H.A., Aung T., Cheng C.Y. Global prevalence of glaucoma and projections of glaucoma burden through 2040: a systematic review and meta-analysis. Ophthalmology. 2014;121:2081–2090. - PubMed
-
- Braunger B.M., Fuchshofer R., Tamm E.R. The aqueous humor outflow pathways in glaucoma: A unifying concept of disease mechanisms and causative treatment. Eur. J. Pharm. Biopharm. 2015;95(Pt B):173–181. - PubMed
-
- Tielsch J.M., Katz J., Sommer A., Quigley H.A., Javitt J.C. Family history and risk of primary open angle glaucoma. The Baltimore Eye Survey. Arch. Ophthalmol. 1994;112:69–73. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
