

Synaptic Properties and Plasticity Mechanisms of Invertebrate Tonic and Phasic Neurons
- PMID: 33391026
- PMCID: PMC7772194
- DOI: 10.3389/fphys.2020.611982
Synaptic Properties and Plasticity Mechanisms of Invertebrate Tonic and Phasic Neurons
Abstract
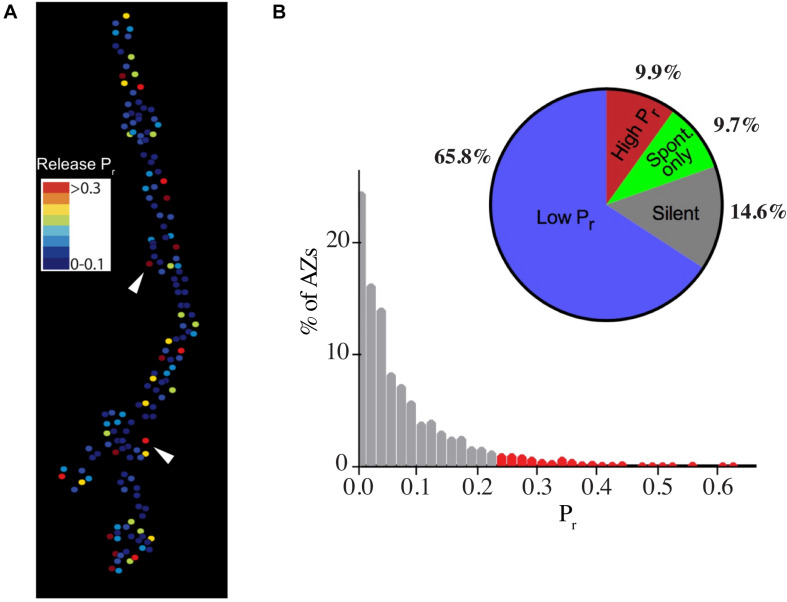
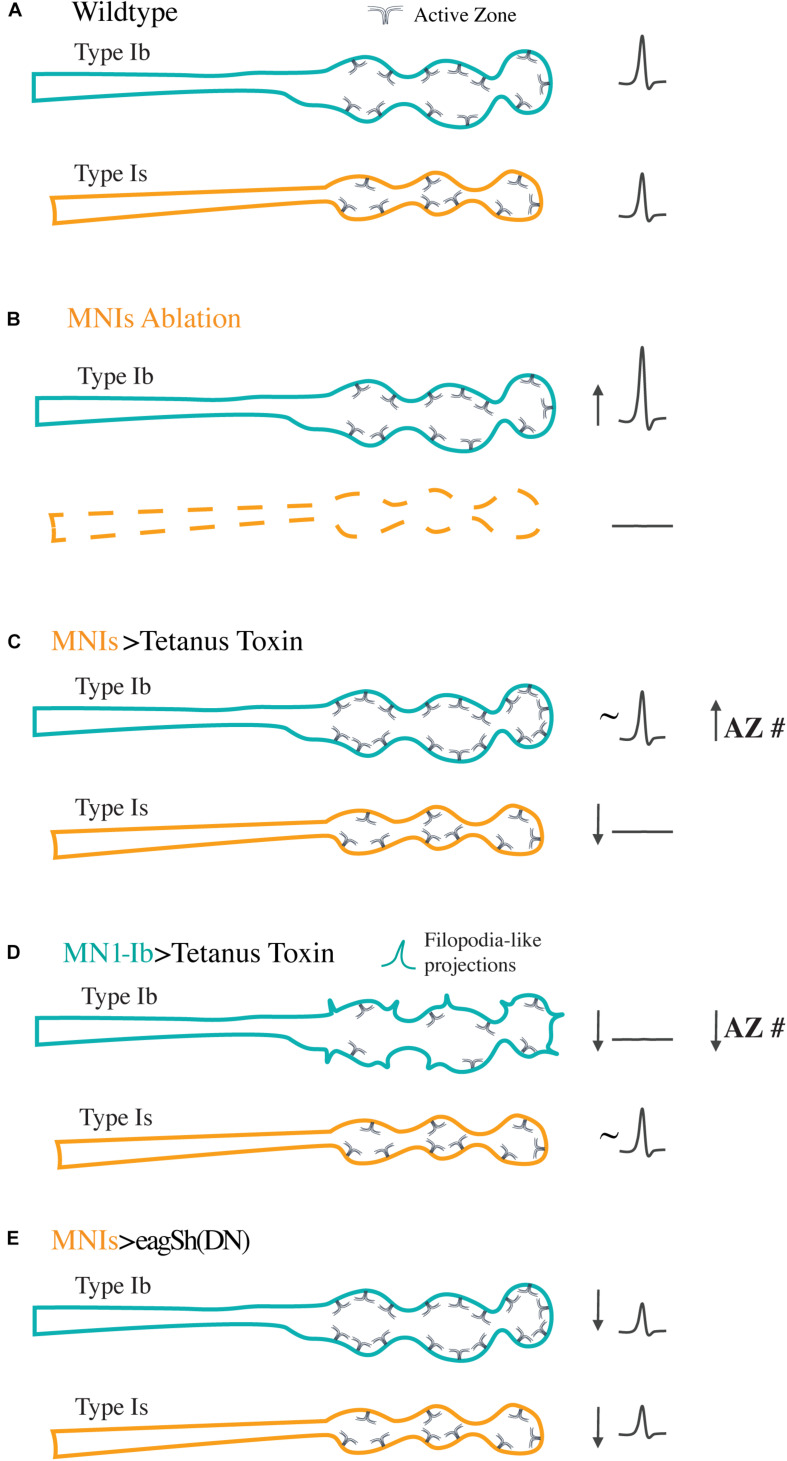
Defining neuronal cell types and their associated biophysical and synaptic diversity has become an important goal in neuroscience as a mechanism to create comprehensive brain cell atlases in the post-genomic age. Beyond broad classification such as neurotransmitter expression, interneuron vs. pyramidal, sensory or motor, the field is still in the early stages of understanding closely related cell types. In both vertebrate and invertebrate nervous systems, one well-described distinction related to firing characteristics and synaptic release properties are tonic and phasic neuronal subtypes. In vertebrates, these classes were defined based on sustained firing responses during stimulation (tonic) vs. transient responses that rapidly adapt (phasic). In crustaceans, the distinction expanded to include synaptic release properties, with tonic motoneurons displaying sustained firing and weaker synapses that undergo short-term facilitation to maintain muscle contraction and posture. In contrast, phasic motoneurons with stronger synapses showed rapid depression and were recruited for short bursts during fast locomotion. Tonic and phasic motoneurons with similarities to those in crustaceans have been characterized in Drosophila, allowing the genetic toolkit associated with this model to be used for dissecting the unique properties and plasticity mechanisms for these neuronal subtypes. This review outlines general properties of invertebrate tonic and phasic motoneurons and highlights recent advances that characterize distinct synaptic and plasticity pathways associated with two closely related glutamatergic neuronal cell types that drive invertebrate locomotion.
Keywords: Drosophila; neuromuscular junction; phasic; synapse; synaptic plasticity; synaptic transmission; tonic.
Copyright © 2020 Aponte-Santiago and Littleton.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures

References
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases