

Extracellular vesicles containing ACE2 efficiently prevent infection by SARS-CoV-2 Spike protein-containing virus
- PMID: 33391636
- PMCID: PMC7769856
- DOI: 10.1002/jev2.12050
Extracellular vesicles containing ACE2 efficiently prevent infection by SARS-CoV-2 Spike protein-containing virus
Abstract
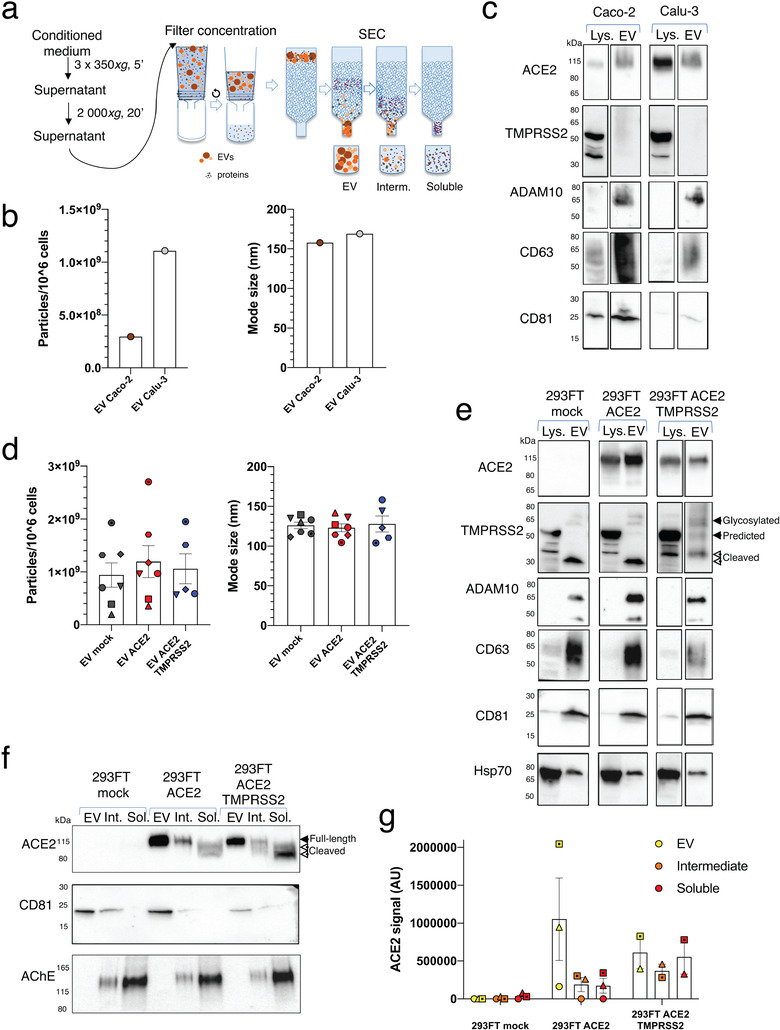
SARS-CoV-2 entry is mediated by binding of the spike protein (S) to the surface receptor ACE2 and subsequent priming by host TMPRSS2 allowing membrane fusion. Here, we produced extracellular vesicles (EVs) exposing ACE2 and demonstrate that ACE2-EVs are efficient decoys for SARS-CoV-2 S protein-containing lentivirus. Reduction of infectivity positively correlates with the level of ACE2, is much more efficient than with soluble ACE2 and further enhanced by the inclusion of TMPRSS2.
Keywords: ACE2; EV therapy; SARS‐CoV‐2; TMPRSS2.
© 2020 The Authors. Journal of Extracellular Vesicles published by Wiley Periodicals, LLC on behalf of the International Society for Extracellular Vesicles.
Figures

References
-
- Afar, D. E. H. , Vivanco, I , Hubert, R. S. , Kuo, J , Chen, E , Saffran, D. C. , Raitano, A. B. , & Jakobovits, A et al. (2001). Catalytic Cleavage of the Androgen‐regulated TMPRSS2 Protease Results in Its Secretion by Prostate and Prostate Cancer Epithelia. Cancer Research, 59, 6015–6022. - PubMed
-
- Börger, V. , Weiss, D. J , Anderson, J. D , Borràs, F. E , Bussolati, B. , Carter, D. R. F , Dominici, M. , Falcón‐Pérez, J. M. , Gimona, M. , Hill, A. F. , Hoffman, A. M. , de Kleijn, D. , Levine, B. L. , Lim, R. , Lötvall, J. , Mitsialis, S. A. , Monguió‐Tortajada, M. , Muraca, M. , Nieuwland, R. … Giebel, B. (2020). International Society for Extracellular Vesicles and International Society for Cell and Gene Therapy statement on extracellular vesicles from mesenchymal stromal cells and other cells: Considerations for potential therapeutic agents to suppress coronavirus disease‐19. Cytotherapy, 22, 482–485 - PMC - PubMed
-
- Cerboni, S. , Jeremiah, N. , Gentili, M. , Gehrmann, U. , Conrad, C. , Stolzenberg, M. C. , Picard, C. , Neven, B. , Fischer, A. , Amigorena, S. , Rieux‐Laucat, F. , & Manel, N. (2017). Intrinsic antiproliferative activity of the innate sensor STING in T lymphocytes. Journal of Experimental Medicine, 214, 1769–1785 - PMC - PubMed
-
- De Carvalho, J. V. , De Castro, R. O. , Da Silva, E Z. M. , Silveira, P. P. , Da Silva‐Januário, M. E. , Arruda, E. , Jamur, M. C. , Oliver, C. , Aguiar, R. S. , & Dasilva, L. L. P. (2014). Nef Neutralizes the Ability of Exosomes from CD4+ T Cells to Act as Decoys during HIV‐1 Infection. Plos One, 9, e113691 - PMC - PubMed
-
- Grzelak, L. , Temmam, S. , Planchais, C. , Demeret, C. , Huon, C. , Guivel‐Benhassine, F. , Staropoli, I. , Chazal, M. , Dufloo, J. , Planas, D. , Buchrieser, J. , Michael Rajah, M. , Robinot, R. , Porrot, F. , Albert, M. , Chen, K.‐Y. , Crescenzo, B. , Donati, F. , Anna, F. , … van der Werf, S. (2020). SARS‐CoV‐2 serological analysis of COVID‐19 hospitalized patients, pauci‐symptomatic individuals and blood donors. medRxiv 2020.04.21.20068858. 10.1101/2020.04.21.20068858 - DOI
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
