

Insights Into the Complexity of Craniofacial Development From a Cellular Perspective
- PMID: 33392208
- PMCID: PMC7775397
- DOI: 10.3389/fcell.2020.620735
Insights Into the Complexity of Craniofacial Development From a Cellular Perspective
Abstract
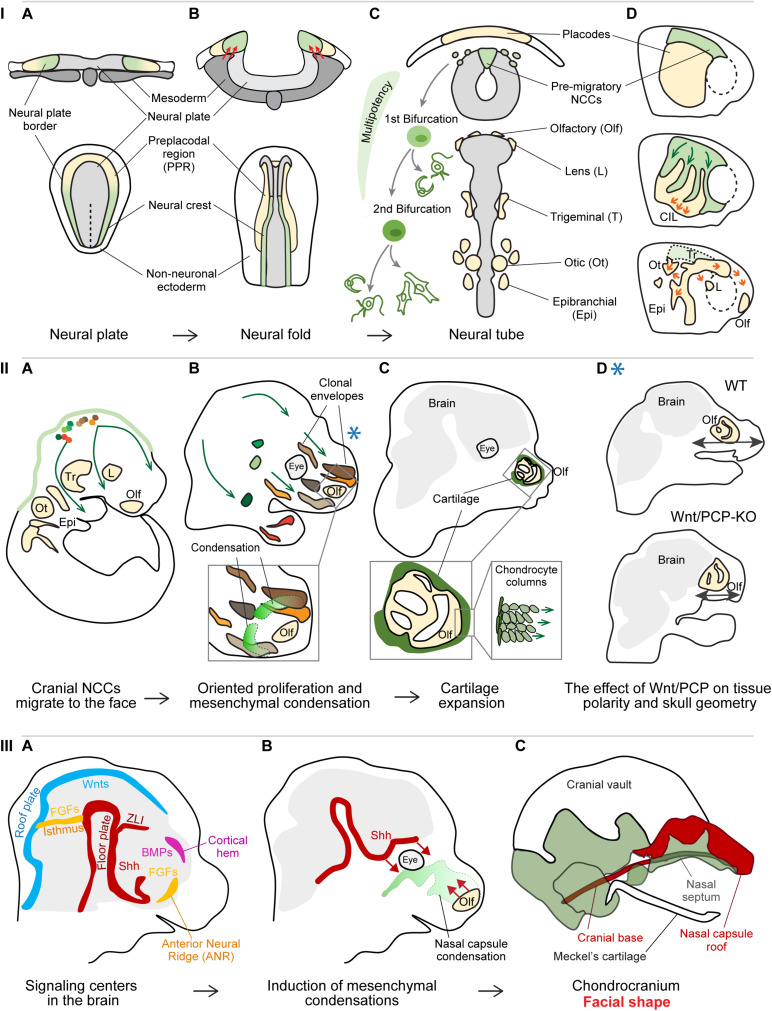
The head represents the most complex part of the body and a distinctive feature of the vertebrate body plan. This intricate structure is assembled during embryonic development in the four-dimensional process of morphogenesis. The head integrates components of the central and peripheral nervous system, sensory organs, muscles, joints, glands, and other specialized tissues in the framework of a complexly shaped skull. The anterior part of the head is referred to as the face, and a broad spectrum of facial shapes across vertebrate species enables different feeding strategies, communication styles, and diverse specialized functions. The face formation starts early during embryonic development and is an enormously complex, multi-step process regulated on a genomic, molecular, and cellular level. In this review, we will discuss recent discoveries that revealed new aspects of facial morphogenesis from the time of the neural crest cell emergence till the formation of the chondrocranium, the primary design of the individual facial shape. We will focus on molecular mechanisms of cell fate specification, the role of individual and collective cell migration, the importance of dynamic and continuous cellular interactions, responses of cells and tissues to generated physical forces, and their morphogenetic outcomes. In the end, we will examine the spatiotemporal activity of signaling centers tightly regulating the release of signals inducing the formation of craniofacial skeletal elements. The existence of these centers and their regulation by enhancers represent one of the core morphogenetic mechanisms and might lay the foundations for intra- and inter-species facial variability.
Keywords: cellular behavior; chondrocranium; craniofacial development; facial shape; mesenchymal condensations; neural crest; signaling centers.
Copyright © 2020 Murillo-Rincón and Kaucka.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures
References
Publication types
LinkOut - more resources
Full Text Sources