

Elucidation and refinement of synthetic receptor mechanisms
- PMID: 33392392
- PMCID: PMC7759213
- DOI: 10.1093/synbio/ysaa017
Elucidation and refinement of synthetic receptor mechanisms
Abstract
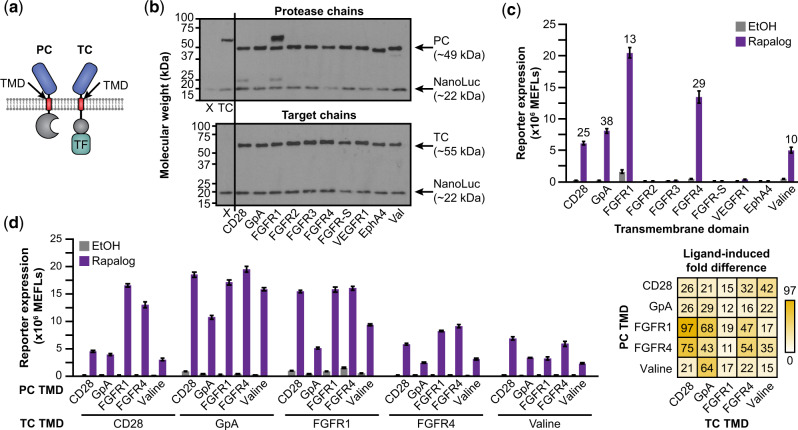
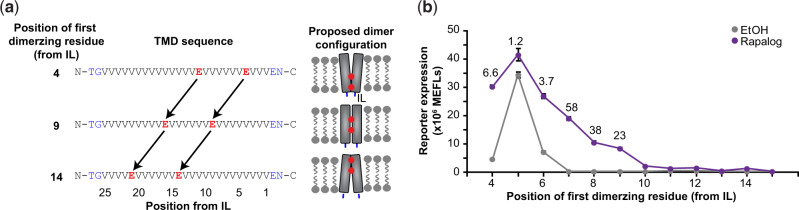
Synthetic receptors are powerful tools for engineering mammalian cell-based devices. These biosensors enable cell-based therapies to perform complex tasks such as regulating therapeutic gene expression in response to sensing physiological cues. Although multiple synthetic receptor systems now exist, many aspects of receptor performance are poorly understood. In general, it would be useful to understand how receptor design choices influence performance characteristics. In this study, we examined the modular extracellular sensor architecture (MESA) and systematically evaluated previously unexamined design choices, yielding substantially improved receptors. A key finding that might extend to other receptor systems is that the choice of transmembrane domain (TMD) is important for generating high-performing receptors. To provide mechanistic insights, we adopted and employed a Förster resonance energy transfer-based assay to elucidate how TMDs affect receptor complex formation and connected these observations to functional performance. To build further insight into these phenomena, we developed a library of new MESA receptors that sense an expanded set of ligands. Based upon these explorations, we conclude that TMDs affect signaling primarily by modulating intracellular domain geometry. Finally, to guide the design of future receptors, we propose general principles for linking design choices to biophysical mechanisms and performance characteristics.
Keywords: biosensor; cell therapy; mammalian synthetic biology; receptor engineering; transmembrane domain.
© The Author(s) 2020. Published by Oxford University Press.
Figures





Similar articles
-
Modular extracellular sensor architecture for engineering mammalian cell-based devices.ACS Synth Biol. 2014 Dec 19;3(12):892-902. doi: 10.1021/sb400128g. Epub 2014 Mar 11. ACS Synth Biol. 2014. PMID: 24611683 Free PMC article.
-
Synthetic Receptors for Sensing Soluble Molecules with Mammalian Cells.Methods Mol Biol. 2021;2312:15-33. doi: 10.1007/978-1-0716-1441-9_2. Methods Mol Biol. 2021. PMID: 34228282
-
Multiplexing Engineered Receptors for Multiparametric Evaluation of Environmental Ligands.ACS Synth Biol. 2017 Nov 17;6(11):2042-2055. doi: 10.1021/acssynbio.6b00279. Epub 2017 Aug 23. ACS Synth Biol. 2017. PMID: 28771312 Free PMC article.
-
The evolution of synthetic receptor systems.Nat Chem Biol. 2022 Mar;18(3):244-255. doi: 10.1038/s41589-021-00926-z. Epub 2022 Jan 20. Nat Chem Biol. 2022. PMID: 35058646 Free PMC article. Review.
-
Membrane receptor activation mechanisms and transmembrane peptide tools to elucidate them.J Biol Chem. 2020 Feb 14;295(7):1792-1814. doi: 10.1074/jbc.REV119.009457. Epub 2019 Dec 25. J Biol Chem. 2020. PMID: 31879273 Free PMC article. Review.
Cited by
-
Post-transcriptional modular synthetic receptors.Nat Chem Biol. 2025 Aug;21(8):1250-1261. doi: 10.1038/s41589-025-01872-w. Epub 2025 Mar 28. Nat Chem Biol. 2025. PMID: 40155716
-
Control of mammalian cell-based devices with genetic programming.Curr Opin Syst Biol. 2021 Dec;28:100372. doi: 10.1016/j.coisb.2021.100372. Epub 2021 Aug 14. Curr Opin Syst Biol. 2021. PMID: 34527830 Free PMC article.
-
Enabling neighbour labelling: using synthetic biology to explore how cells influence their neighbours.Development. 2024 Jan 1;151(1):dev201955. doi: 10.1242/dev.201955. Epub 2024 Jan 2. Development. 2024. PMID: 38165174 Free PMC article. Review.
-
Model-guided design of mammalian genetic programs.Sci Adv. 2021 Feb 19;7(8):eabe9375. doi: 10.1126/sciadv.abe9375. Print 2021 Feb. Sci Adv. 2021. PMID: 33608279 Free PMC article.
-
Building Synthetic Biosensors Using Red Blood Cell Proteins.ACS Synth Biol. 2024 Apr 19;13(4):1273-1289. doi: 10.1021/acssynbio.3c00754. Epub 2024 Mar 27. ACS Synth Biol. 2024. PMID: 38536408 Free PMC article.
References
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials