

Mitigating oxygen stress enhances aged mouse hematopoietic stem cell numbers and function
- PMID: 33393491
- PMCID: PMC7773345
- DOI: 10.1172/JCI140177
Mitigating oxygen stress enhances aged mouse hematopoietic stem cell numbers and function
Abstract
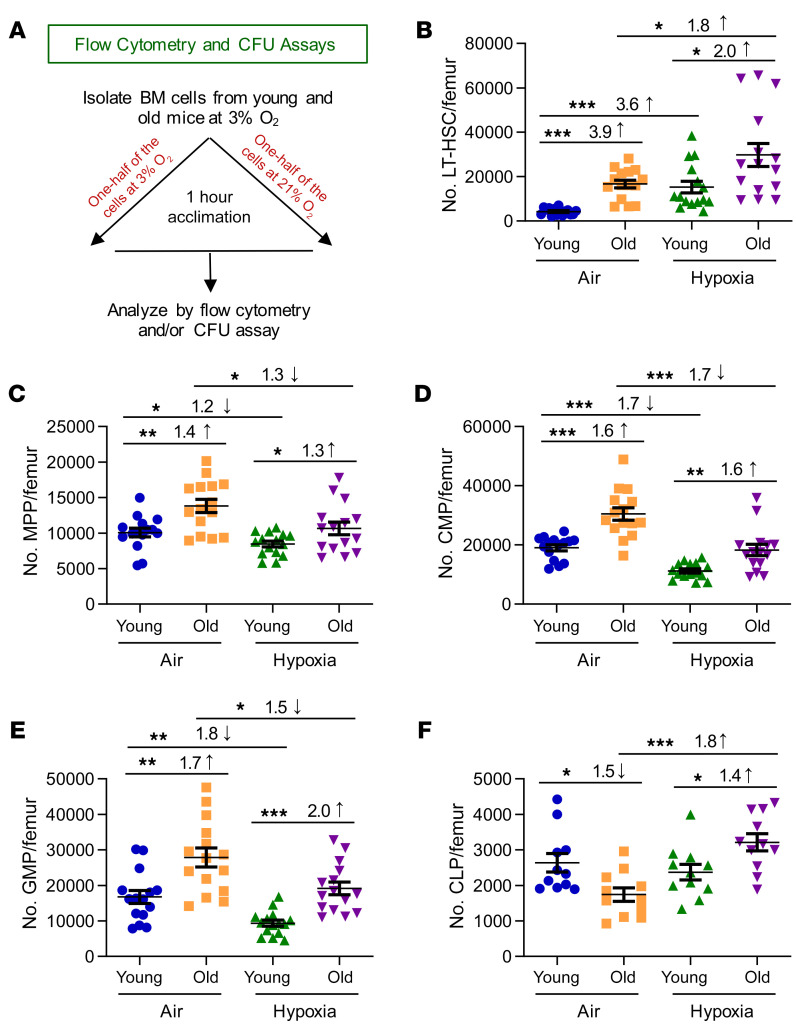
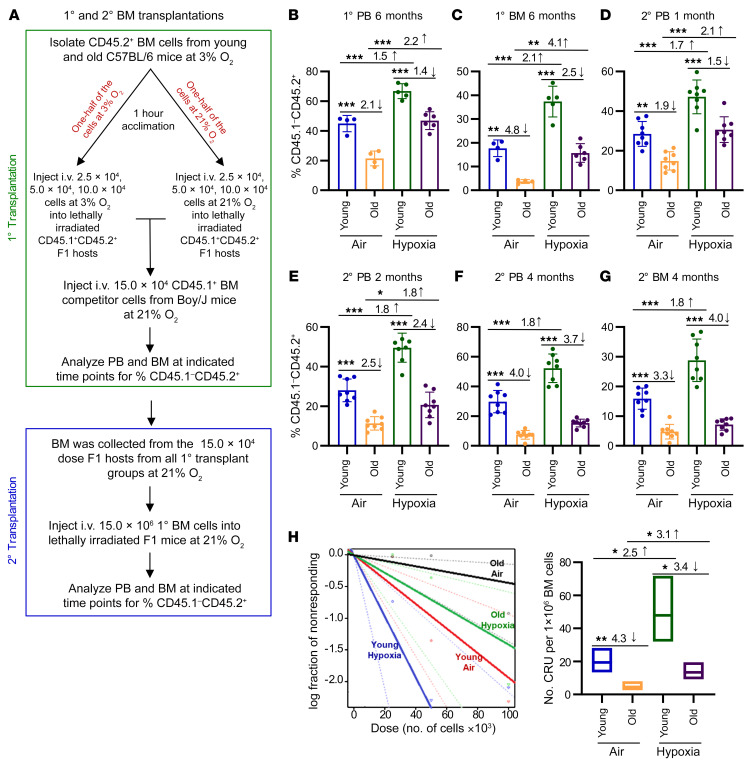
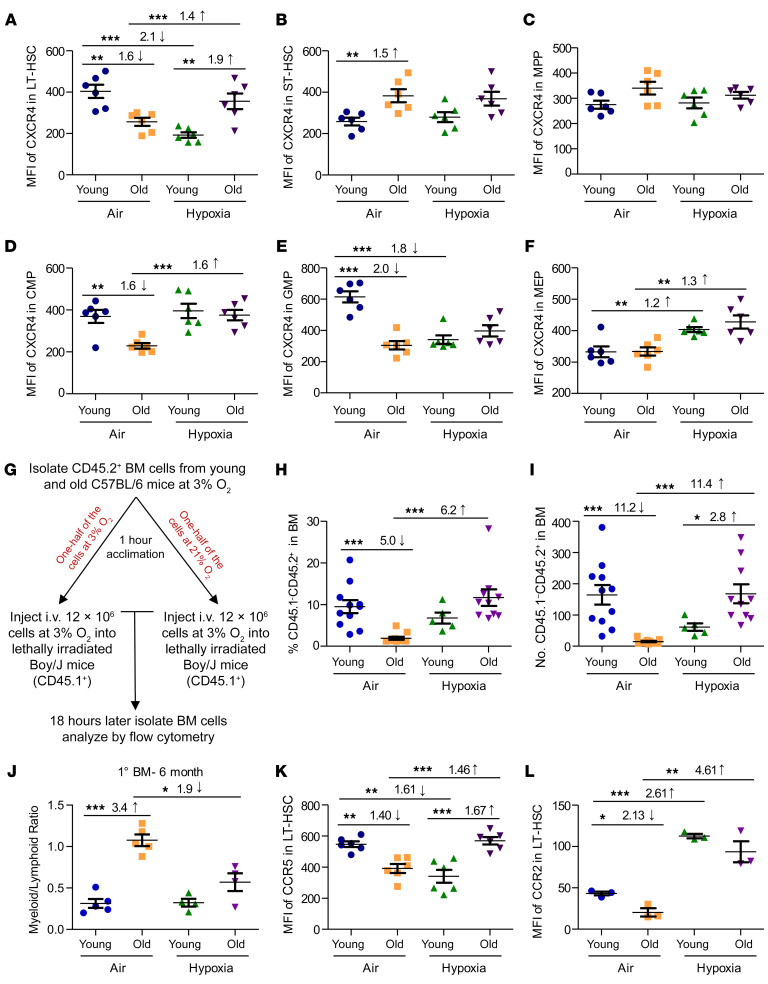
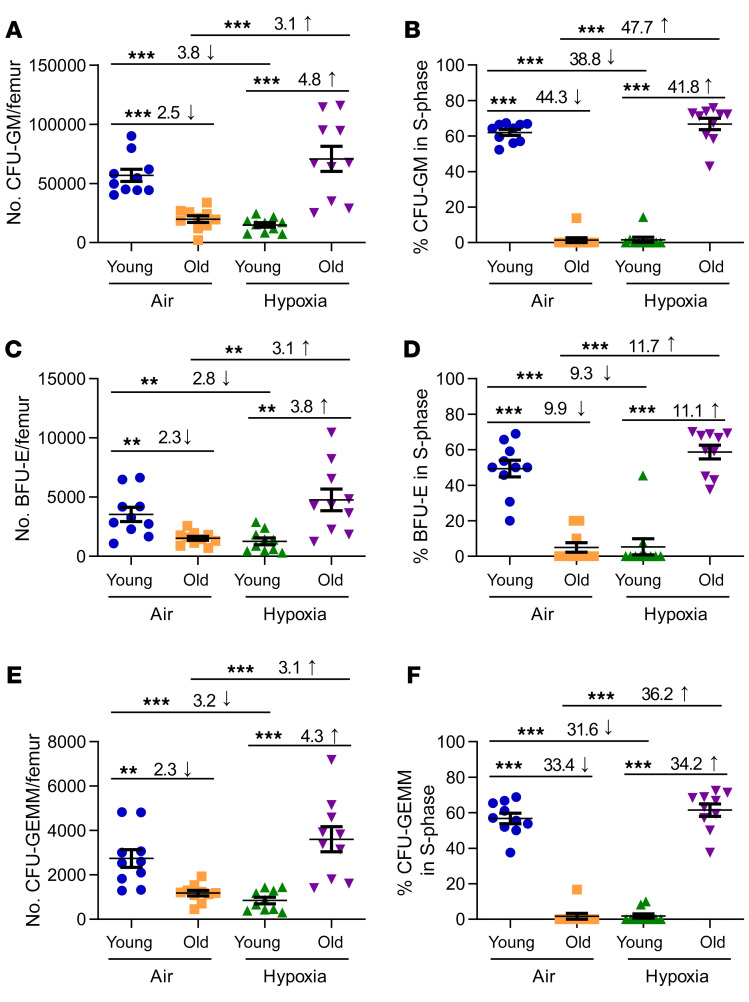
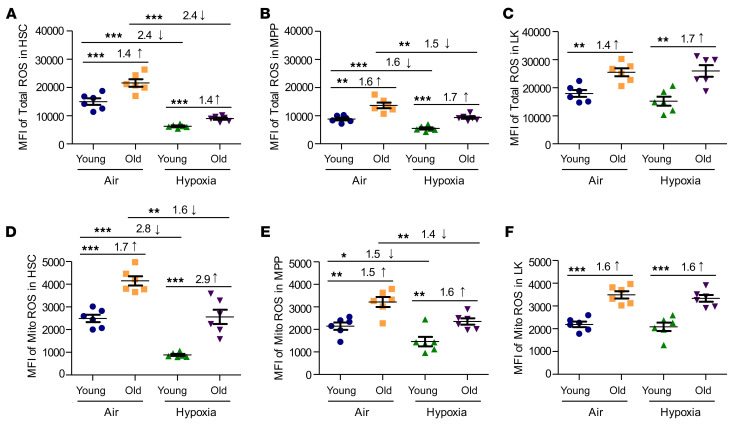
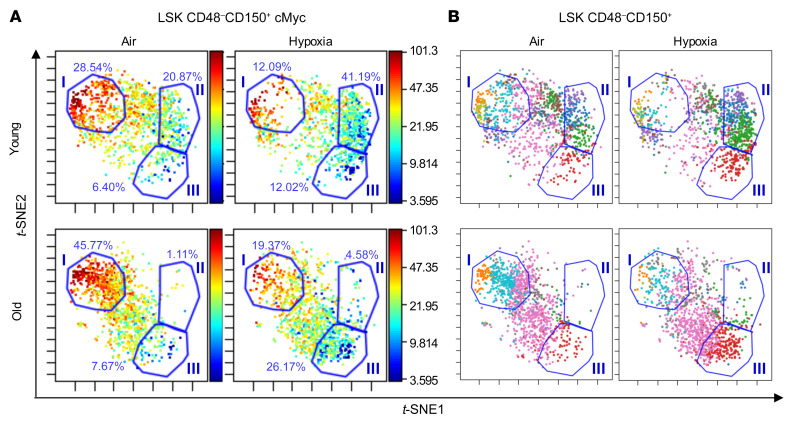
Bone marrow (BM) hematopoietic stem cells (HSCs) become dysfunctional during aging (i.e., they are increased in number but have an overall reduction in long-term repopulation potential and increased myeloid differentiation) compared with young HSCs, suggesting limited use of old donor BM cells for hematopoietic cell transplantation (HCT). BM cells reside in an in vivo hypoxic environment yet are evaluated after collection and processing in ambient air. We detected an increase in the number of both young and aged mouse BM HSCs collected and processed in 3% O2 compared with the number of young BM HSCs collected and processed in ambient air (~21% O2). Aged BM collected and processed under hypoxic conditions demonstrated enhanced engraftment capability during competitive transplantation analysis and contained more functional HSCs as determined by limiting dilution analysis. Importantly, the myeloid-to-lymphoid differentiation ratio of aged BM collected in 3% O2 was similar to that detected in young BM collected in ambient air or hypoxic conditions, consistent with the increased number of common lymphoid progenitors following collection under hypoxia. Enhanced functional activity and differentiation of old BM collected and processed in hypoxia correlated with reduced "stress" associated with ambient air BM collection and suggests that aged BM may be better and more efficiently used for HCT if collected and processed under hypoxia so that it is never exposed to ambient air O2.
Keywords: Aging; Bone marrow transplantation; Hematology; Hematopoietic stem cells; hypoxia.
Conflict of interest statement
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
