

Cyclin-dependent Kinase 1 and Aurora Kinase choreograph mitotic storage and redistribution of a growth factor receptor
- PMID: 33395410
- PMCID: PMC7808676
- DOI: 10.1371/journal.pbio.3001029
Cyclin-dependent Kinase 1 and Aurora Kinase choreograph mitotic storage and redistribution of a growth factor receptor
Abstract
Endosomal trafficking of receptors and associated proteins plays a critical role in signal processing. Until recently, it was thought that trafficking was shut down during cell division. Thus, remarkably, the regulation of trafficking during division remains poorly characterized. Here we delineate the role of mitotic kinases in receptor trafficking during asymmetric division. Targeted perturbations reveal that Cyclin-dependent Kinase 1 (CDK1) and Aurora Kinase promote storage of Fibroblast Growth Factor Receptors (FGFRs) by suppressing endosomal degradation and recycling pathways. As cells progress through metaphase, loss of CDK1 activity permits differential degradation and targeted recycling of stored receptors, leading to asymmetric induction. Mitotic receptor storage, as delineated in this study, may facilitate rapid reestablishment of signaling competence in nascent daughter cells. However, mutations that limit or enhance the release of stored signaling components could alter daughter cell fate or behavior thereby promoting oncogenesis.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures


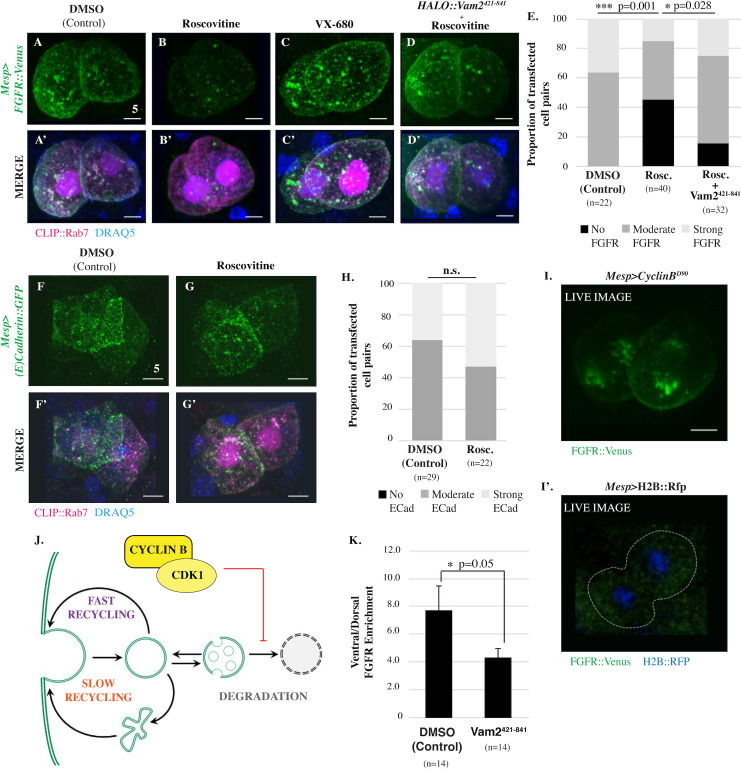
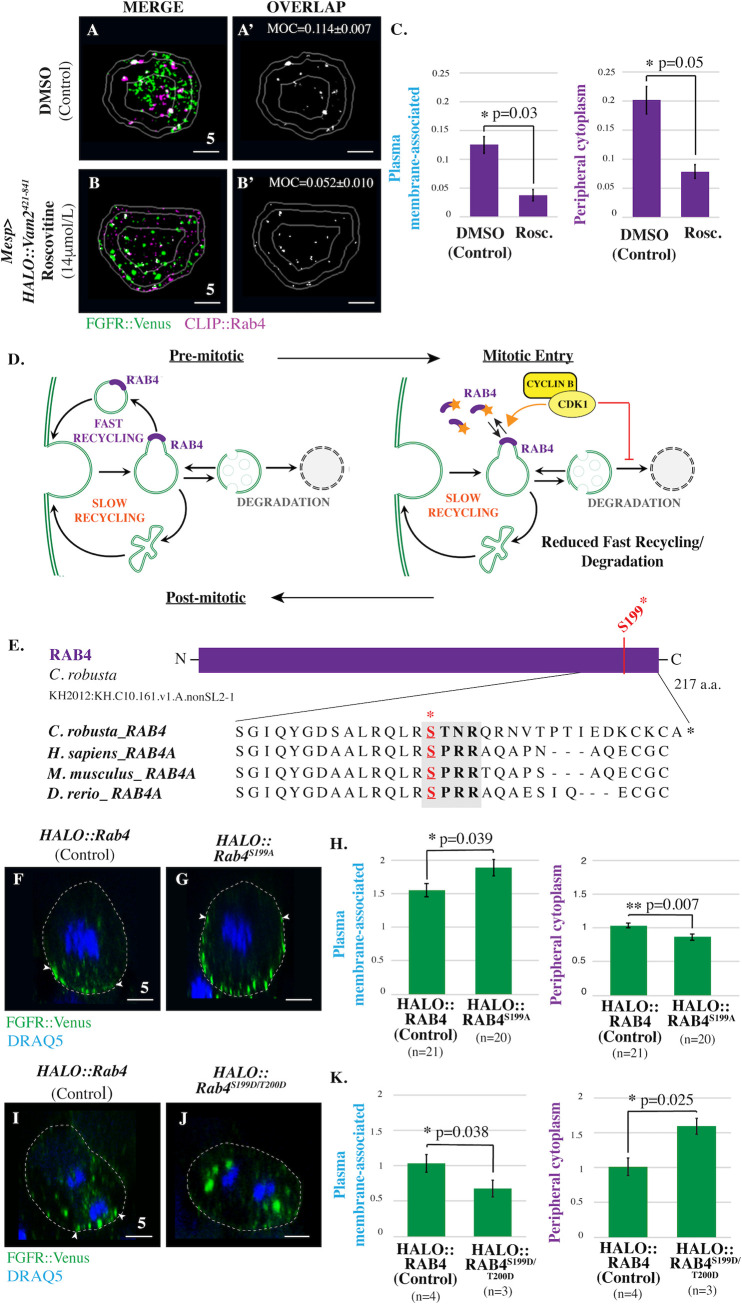

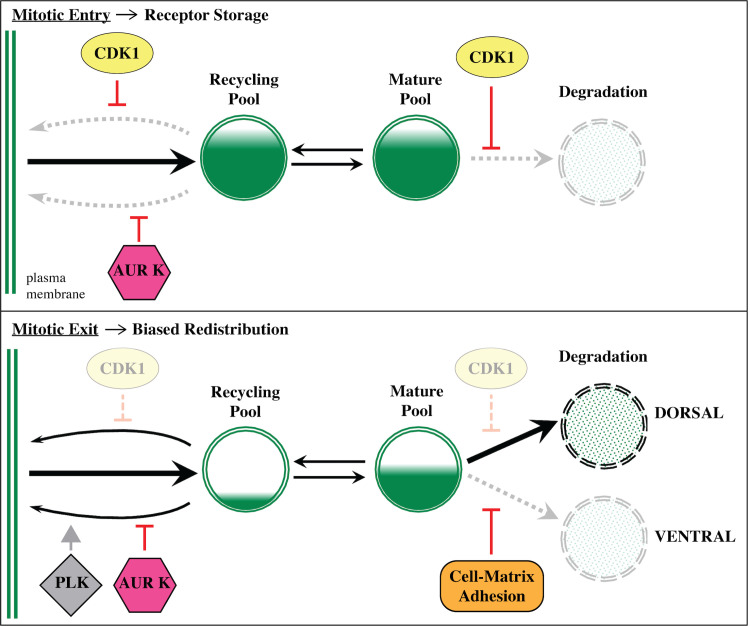
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
