

Off-target effects of RNAi correlate with the mismatch rate between dsRNA and non-target mRNA
- PMID: 33397184
- PMCID: PMC8583100
- DOI: 10.1080/15476286.2020.1868680
Off-target effects of RNAi correlate with the mismatch rate between dsRNA and non-target mRNA
Abstract
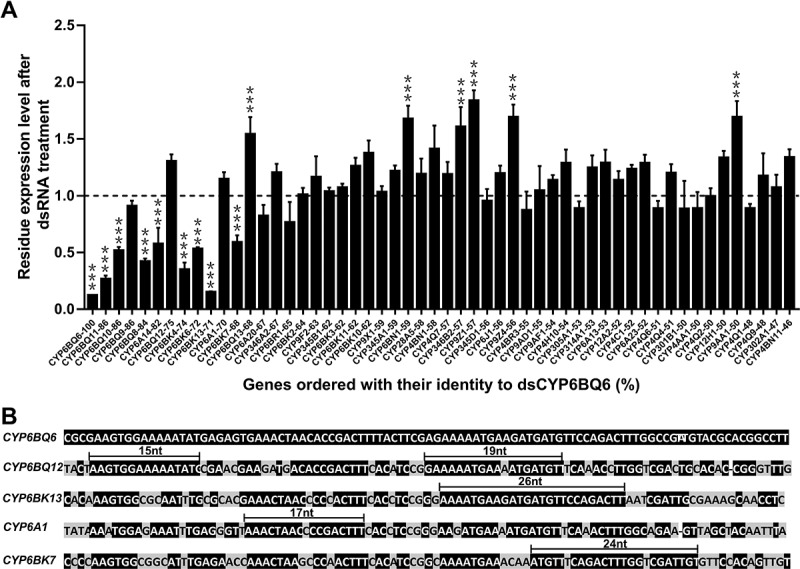
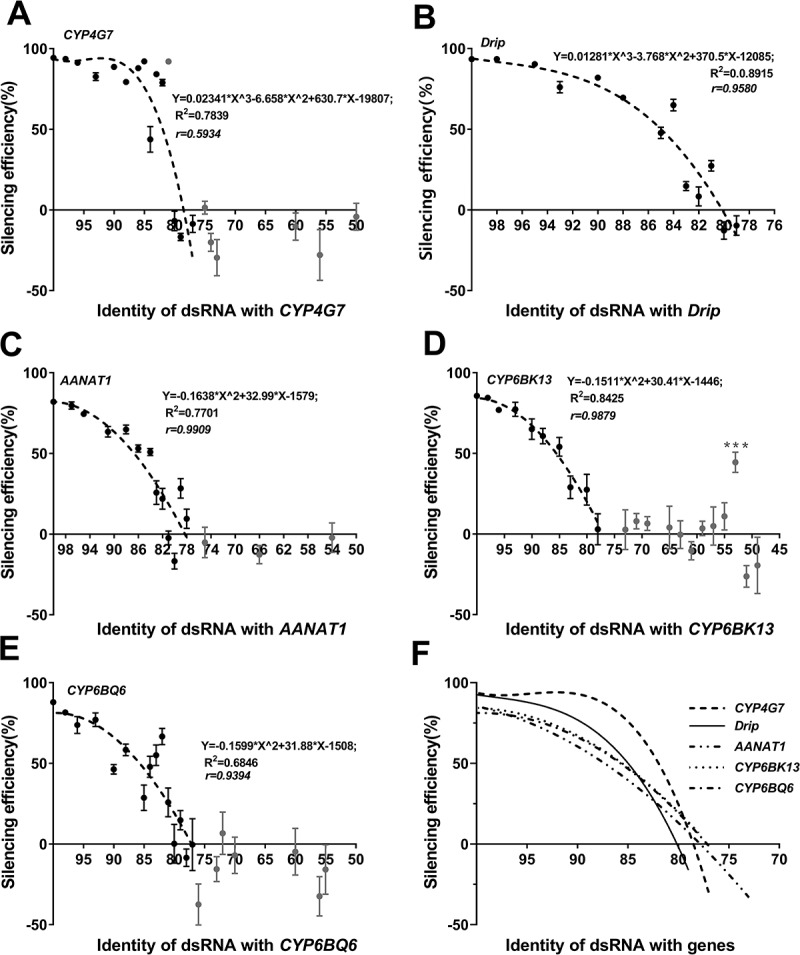
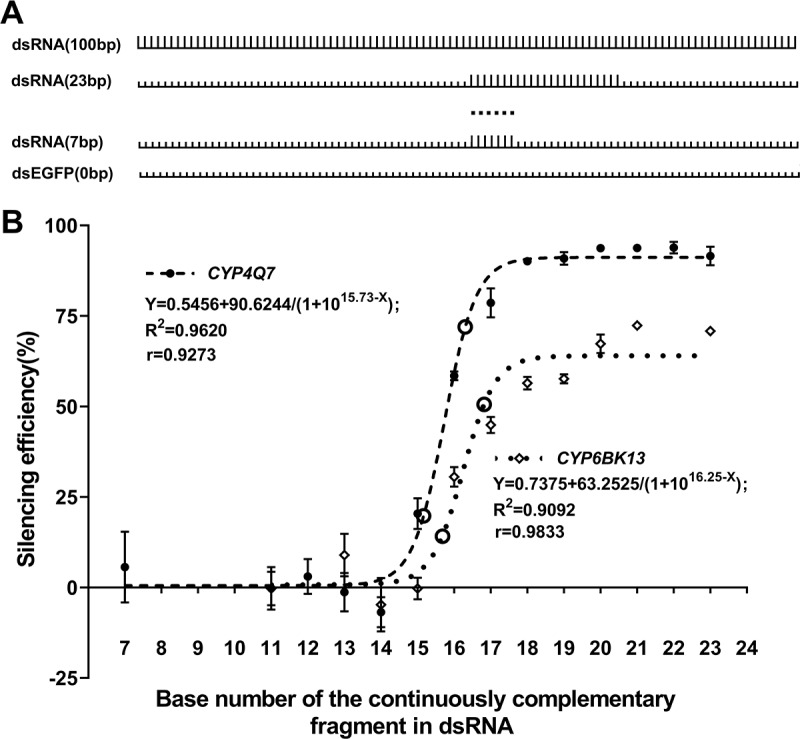
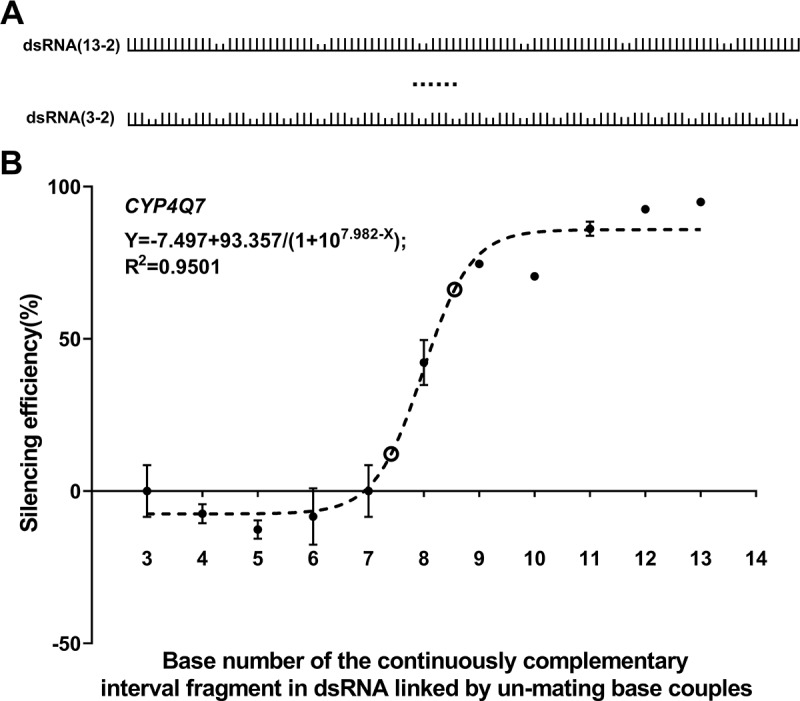
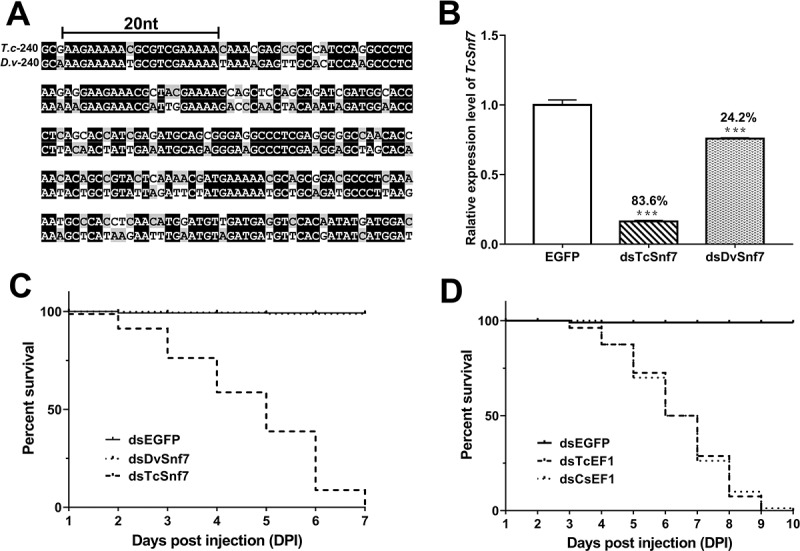
RNAi is a potent technique for the knockdown of target genes. However, its potential off-target effects limit the widespread applications in both reverse genetic analysis and genetic manipulation. Previous efforts have uncovered rules underlying specificity of siRNA-based silencing, which has broad applications in humans, but the basis for specificity of dsRNAs, which are better suited for use as insecticides, is poorly understood. Here, we investigated the rules governing dsRNA specificity. Mutational analyses showed that dsRNAs with >80% sequence identity with target genes triggered RNAi efficiently. dsRNAs with ≥16 bp segments of perfectly matched sequence or >26 bp segments of almost perfectly matched sequence with one or two mismatches scarcely distributed (single mismatches inserted between ≥5 bp matching segments or mismatched couplets inserted between ≥8 bp matching segments) also able to trigger RNAi. Using these parameters to predict off-target risk, dsRNAs can be designed to optimize specificity and efficiency, paving the way to the widespread, rational application of RNAi in pest control.
Keywords: RNA interference; RNAi efficiency; dsRNA specificity; off-target effect; risk assessment; sequence identity.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources