

Inhibition of interferon-signalling halts cancer-associated fibroblast-dependent protection of breast cancer cells from chemotherapy
- PMID: 33398063
- PMCID: PMC7960738
- DOI: 10.1038/s41416-020-01226-4
Inhibition of interferon-signalling halts cancer-associated fibroblast-dependent protection of breast cancer cells from chemotherapy
Abstract
Background: Triple negative breast cancers (TNBC) have poor prognoses despite aggressive treatment with cytotoxic chemotherapy. Cancer-associated fibroblasts (CAFs) are prominent in tumour stroma. Our hypothesis was that CAFs modulate chemotherapy sensitivity.
Methods: TNBC cells and breast fibroblasts were cultured; survival after chemotherapeutics was assessed using luciferase or clonogenic assays. Signalling was investigated using transcriptomics, reporters, recombinant proteins and blocking antibodies. Clinical relevance was investigated using immunohistochemistry.
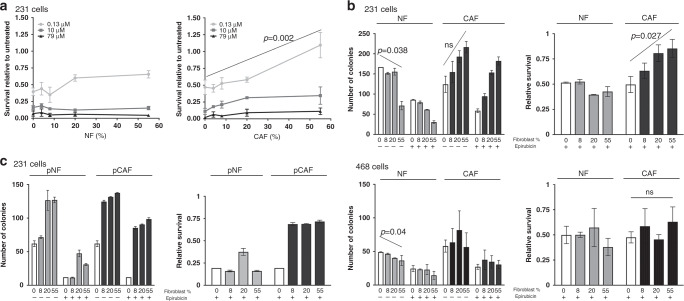
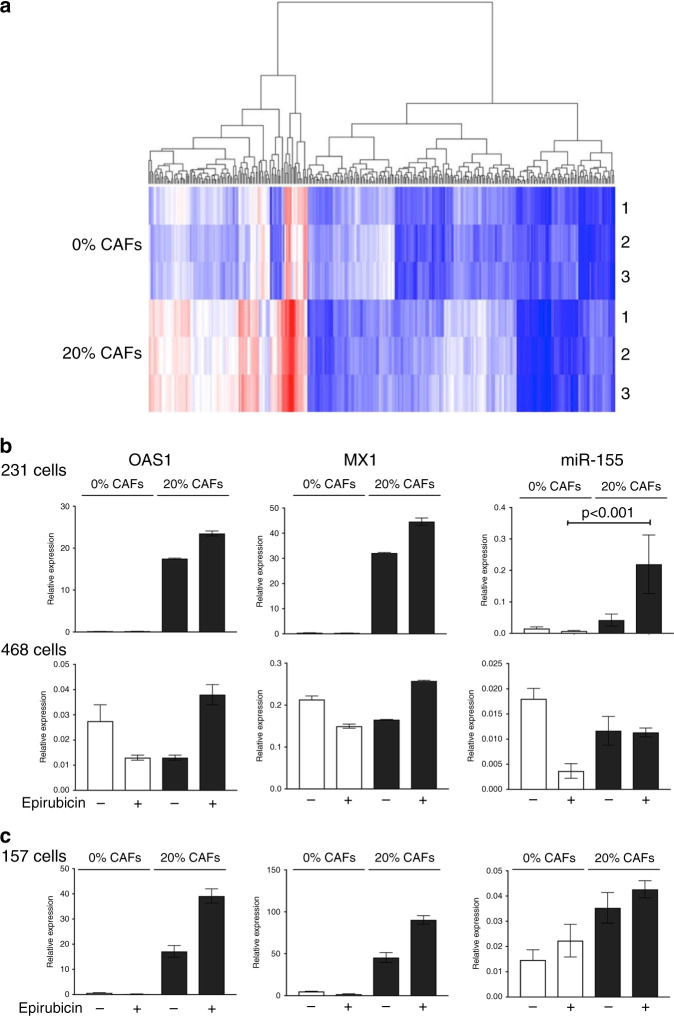
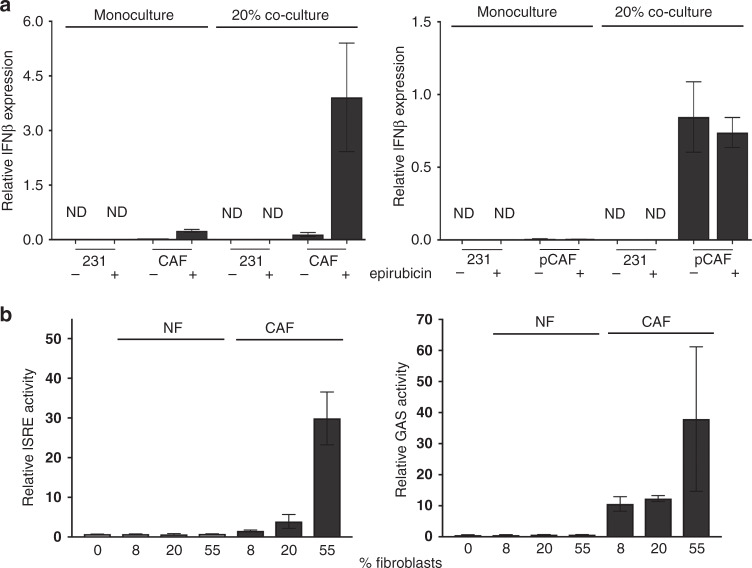
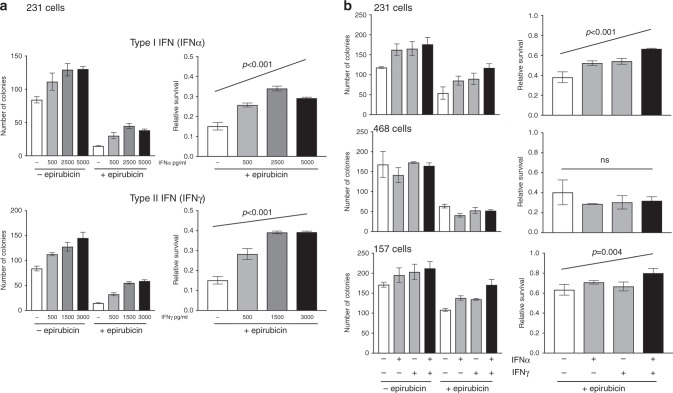
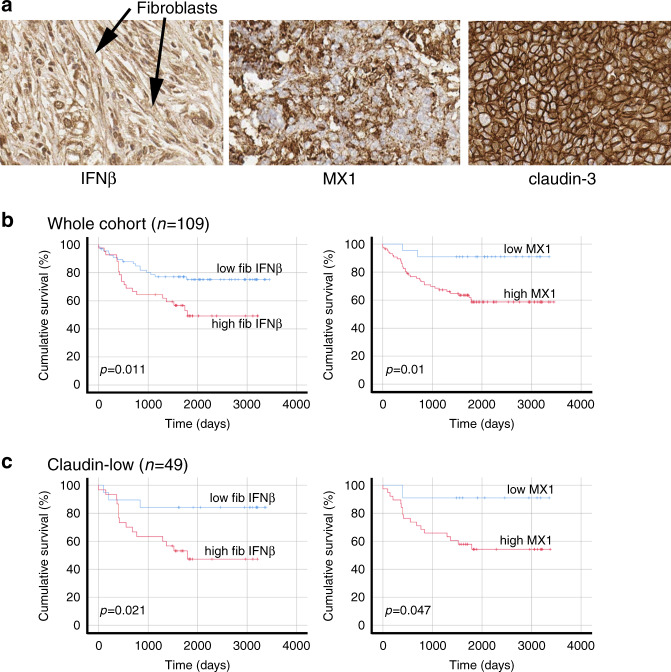
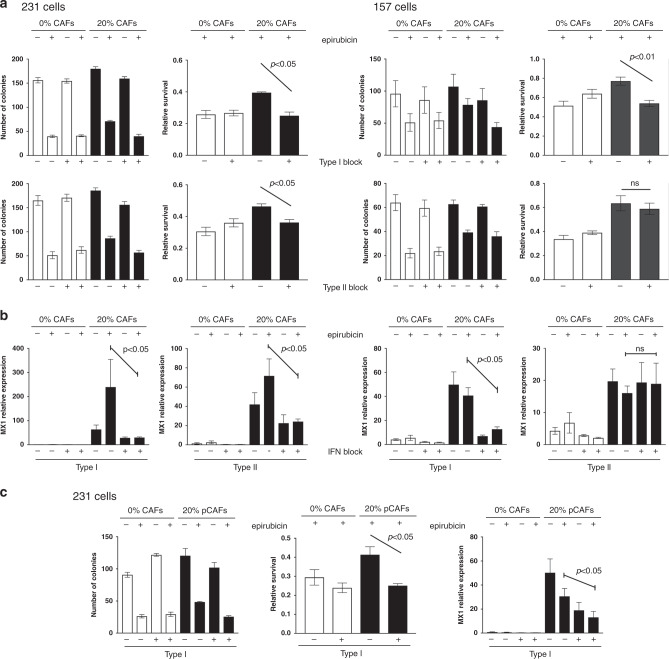
Results: Breast CAFs dose-dependently protected TNBC cell lines MDA-MB-231 and MDA-MB-157, but not MDA-MB-468s, from chemotherapy. CAF-induced protection was associated with interferon (IFN) activation. CAFs were induced to express IFNβ1 by chemotherapy and TNBC co-culture, leading to paracrine activation in cancer cells. Recombinant IFNs were sufficient to protect MDA-MB-231 and MDA-MB-157 but not MDA-MB-468 cells. In TNBC patients, IFNβ1 expression in CAFs correlated with cancer cell expression of MX1, a marker of activated IFN signalling. High expression of IFNβ1 (CAFs) or MX1 (tumour cells) correlated with reduced survival after chemotherapy, especially in claudin-low tumours (which MDA-MB-231 and MDA-MB-157 cells represent). Antibodies that block IFN receptors reduced CAF-dependent chemoprotection.
Conclusions: CAF-induced activation of IFN signalling in claudin-low TNBCs results in chemoresistance. Inhibition of this pathway represents a novel method to improve breast cancer outcomes.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
