

Mechanisms controlling pancreatic islet cell function in insulin secretion
- PMID: 33398164
- PMCID: PMC8115730
- DOI: 10.1038/s41580-020-00317-7
Mechanisms controlling pancreatic islet cell function in insulin secretion
Abstract
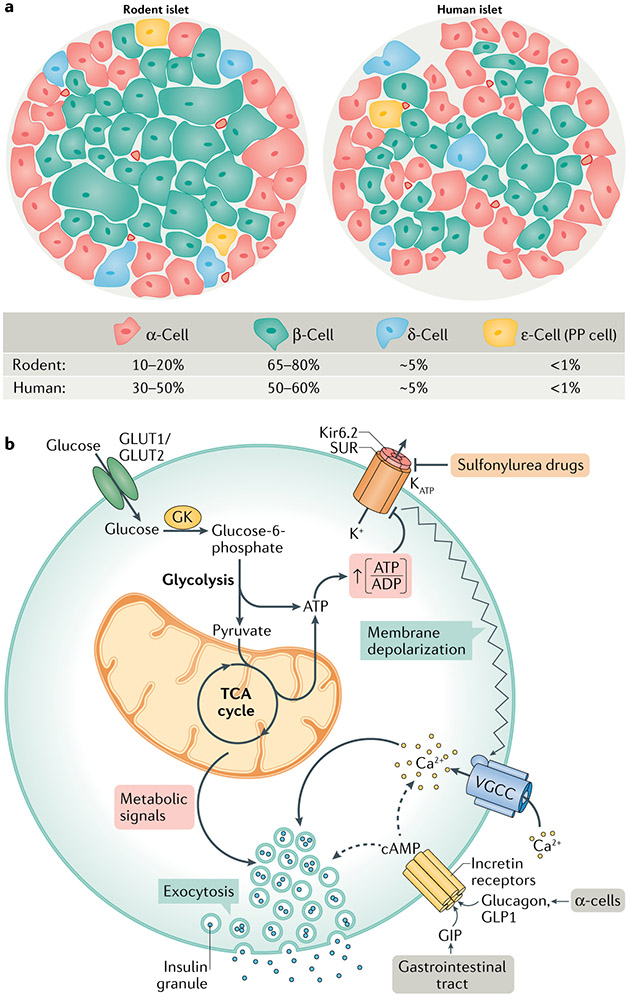
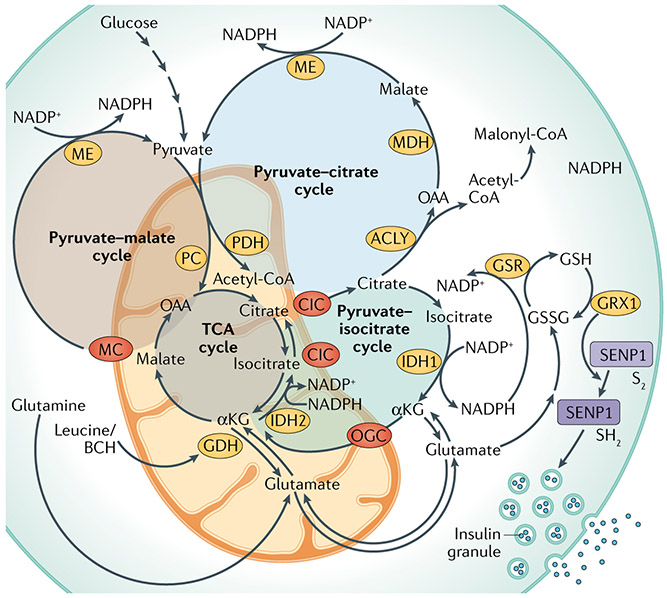
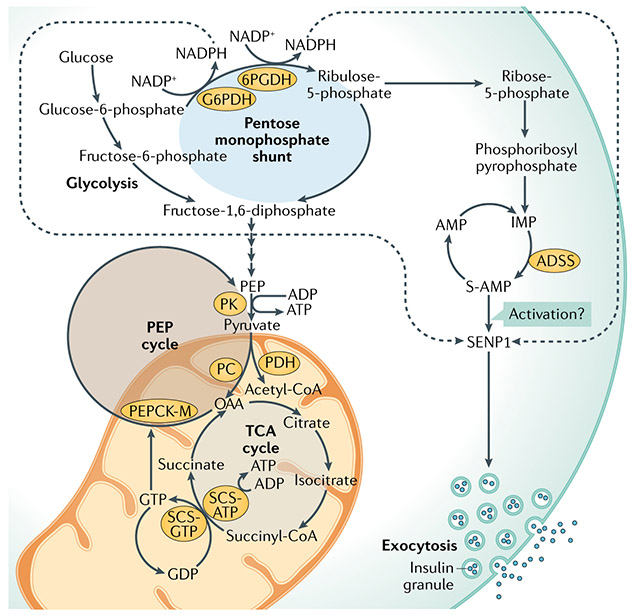
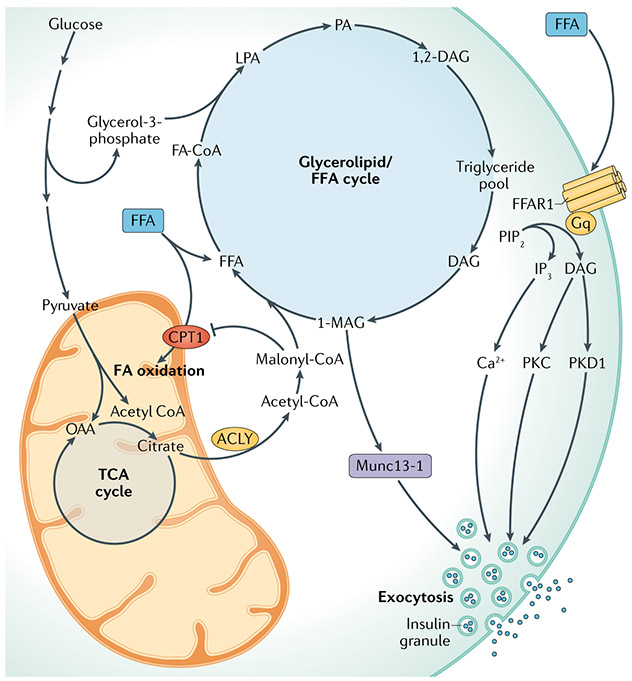
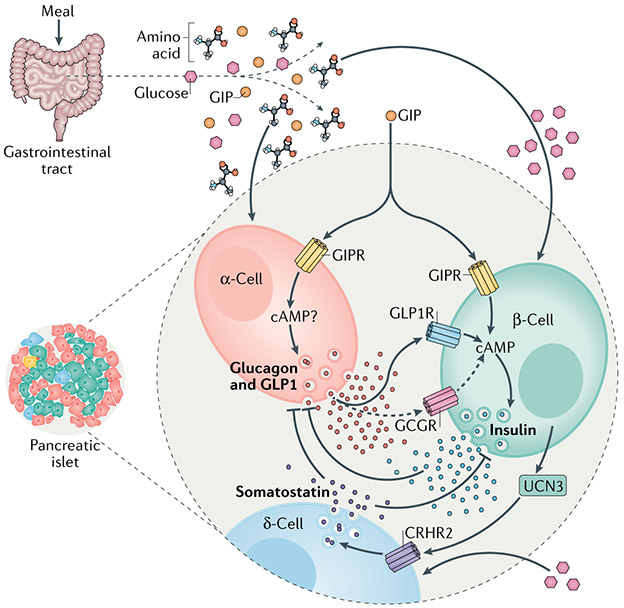
Metabolic homeostasis in mammals is tightly regulated by the complementary actions of insulin and glucagon. The secretion of these hormones from pancreatic β-cells and α-cells, respectively, is controlled by metabolic, endocrine, and paracrine regulatory mechanisms and is essential for the control of blood levels of glucose. The deregulation of these mechanisms leads to various pathologies, most notably type 2 diabetes, which is driven by the combined lesions of impaired insulin action and a loss of the normal insulin secretion response to glucose. Glucose stimulates insulin secretion from β-cells in a bi-modal fashion, and new insights about the underlying mechanisms, particularly relating to the second or amplifying phase of this secretory response, have been recently gained. Other recent work highlights the importance of α-cell-produced proglucagon-derived peptides, incretin hormones from the gastrointestinal tract and other dietary components, including certain amino acids and fatty acids, in priming and potentiation of the β-cell glucose response. These advances provide a new perspective for the understanding of the β-cell failure that triggers type 2 diabetes.
Figures
References
-
- Saito K, Takahashi T, Yaginuma N & Iwama N Islet morphometry in the diabetic pancreas of man. Tohoku J. Exp. Med 125, 185–197 (1978). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
