

Hyaluronidase Impairs Neutrophil Function and Promotes Group B Streptococcus Invasion and Preterm Labor in Nonhuman Primates
- PMID: 33402537
- PMCID: PMC8545101
- DOI: 10.1128/mBio.03115-20
Hyaluronidase Impairs Neutrophil Function and Promotes Group B Streptococcus Invasion and Preterm Labor in Nonhuman Primates
Abstract
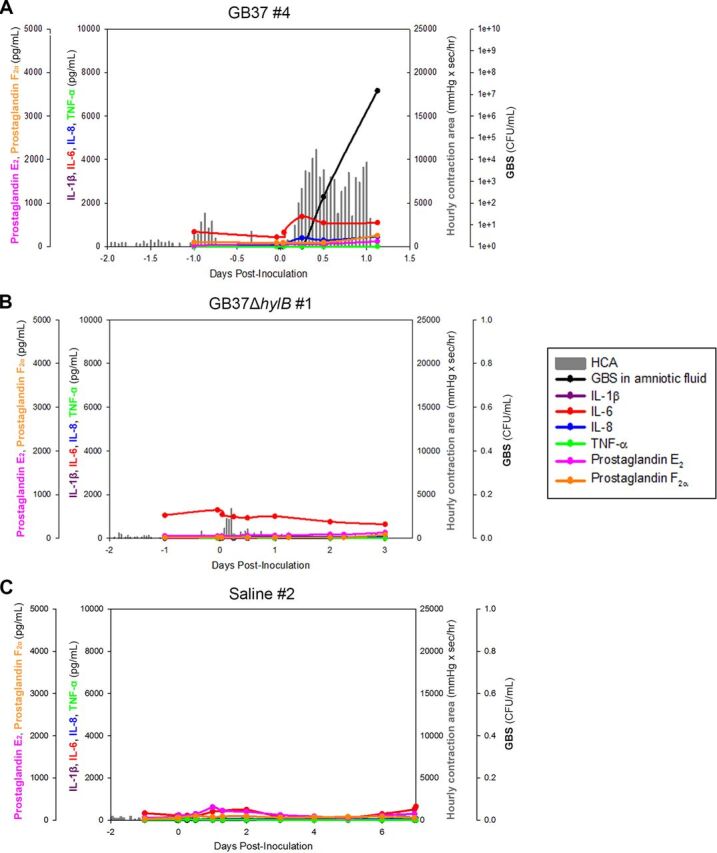
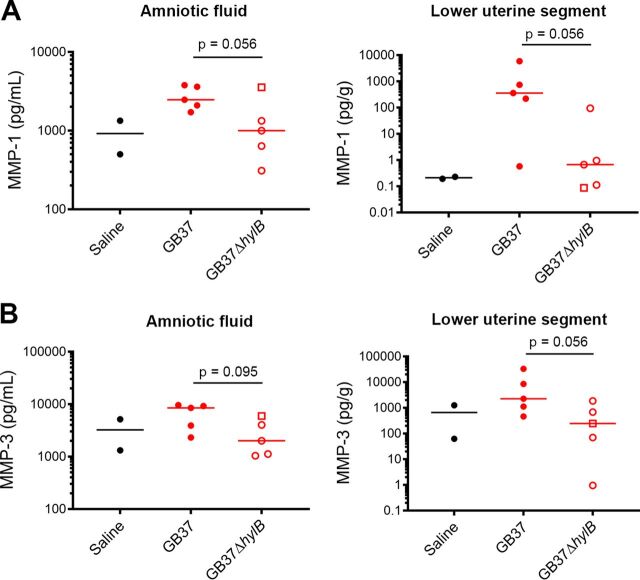
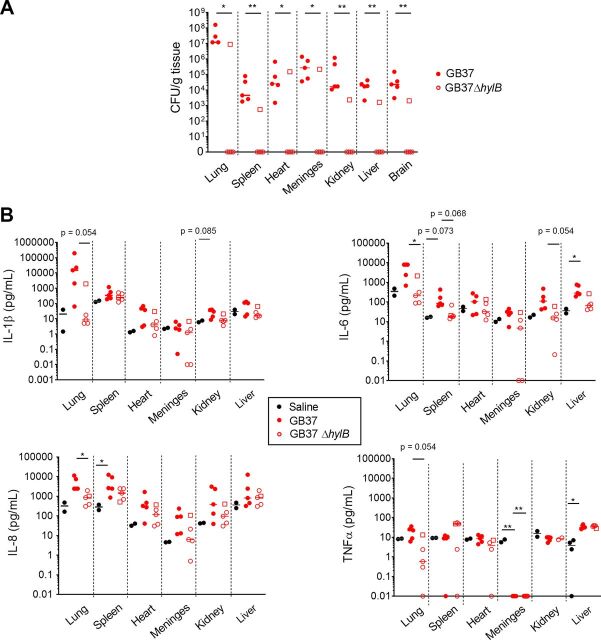
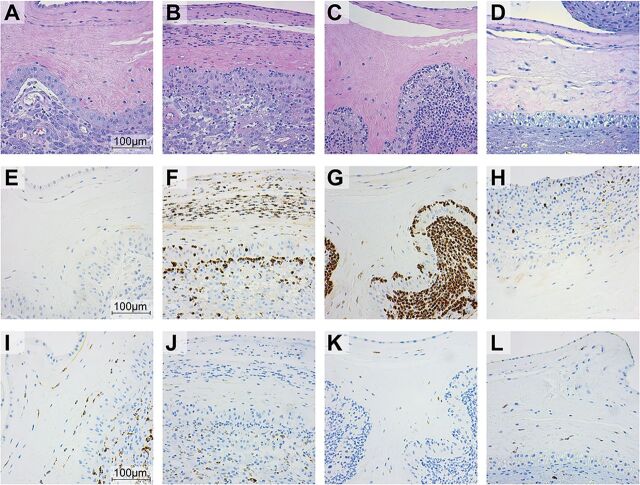
Invasive bacterial infections during pregnancy are a major risk factor for preterm birth, stillbirth, and fetal injury. Group B streptococci (GBS) are Gram-positive bacteria that asymptomatically colonize the lower genital tract but infect the amniotic fluid and induce preterm birth or stillbirth. Experimental models that closely emulate human pregnancy are pivotal for the development of successful strategies to prevent these adverse pregnancy outcomes. Using a unique nonhuman primate model that mimics human pregnancy and informs temporal events surrounding amniotic cavity invasion and preterm labor, we show that the animals inoculated with hyaluronidase (HylB)-expressing GBS consistently exhibited microbial invasion into the amniotic cavity, fetal bacteremia, and preterm labor. Although delayed cytokine responses were observed at the maternal-fetal interface, increased prostaglandin and matrix metalloproteinase levels in these animals likely mediated preterm labor. HylB-proficient GBS dampened reactive oxygen species production and exhibited increased resistance to neutrophils compared to an isogenic mutant. Together, these findings demonstrate how a bacterial enzyme promotes GBS amniotic cavity invasion and preterm labor in a model that closely resembles human pregnancy.IMPORTANCE Group B streptococci (GBS) are bacteria that commonly reside in the female lower genital tract as asymptomatic members of the microbiota. However, during pregnancy, GBS can infect tissues at the maternal-fetal interface, leading to preterm birth, stillbirth, or fetal injury. Understanding how GBS evade host defenses during pregnancy is key to developing improved preventive therapies for these adverse outcomes. In this study, we used a unique nonhuman primate model to show that an enzyme secreted by GBS, hyaluronidase (HylB) promotes bacterial invasion into the amniotic cavity and fetus. Although delayed immune responses were seen at the maternal-fetal interface, animals infected with hyaluronidase-expressing GBS exhibited premature cervical ripening and preterm labor. These observations reveal that HylB is a crucial GBS virulence factor that promotes bacterial invasion and preterm labor in a pregnancy model that closely emulates human pregnancy. Therefore, hyaluronidase inhibitors may be useful in therapeutic strategies against ascending GBS infection.
Keywords: group B streptococcus; hyaluronidase; immune evasion; neutrophils; pregnancy; preterm labor.
Copyright © 2021 Coleman et al.
Figures
Similar articles
-
Bacterial Hyaluronidase Promotes Ascending GBS Infection and Preterm Birth.mBio. 2016 Jun 28;7(3):e00781-16. doi: 10.1128/mBio.00781-16. mBio. 2016. PMID: 27353757 Free PMC article.
-
Amniotic fluid interleukin 6 and interleukin 8 are superior predictors of fetal lung injury compared with maternal or fetal plasma cytokines or placental histopathology in a nonhuman primate model.Am J Obstet Gynecol. 2021 Jul;225(1):89.e1-89.e16. doi: 10.1016/j.ajog.2020.12.1214. Epub 2021 Jan 4. Am J Obstet Gynecol. 2021. PMID: 33412130 Free PMC article.
-
A Broad Spectrum Chemokine Inhibitor Prevents Preterm Labor but Not Microbial Invasion of the Amniotic Cavity or Neonatal Morbidity in a Non-human Primate Model.Front Immunol. 2020 Apr 30;11:770. doi: 10.3389/fimmu.2020.00770. eCollection 2020. Front Immunol. 2020. PMID: 32425945 Free PMC article.
-
Mechanisms of group B Streptococcus-mediated preterm birth: lessons learnt from animal models.Reprod Fertil. 2022 Jun 7;3(3):R109-R120. doi: 10.1530/RAF-21-0105. eCollection 2022 Jul 1. Reprod Fertil. 2022. PMID: 35794927 Free PMC article. Review.
-
Perinatal Group B Streptococcal Infections: Virulence Factors, Immunity, and Prevention Strategies.Trends Microbiol. 2017 Nov;25(11):919-931. doi: 10.1016/j.tim.2017.05.013. Epub 2017 Jun 17. Trends Microbiol. 2017. PMID: 28633864 Free PMC article. Review.
Cited by
-
Spatial profiling of the placental chorioamniotic membranes reveals upregulation of immune checkpoint proteins during Group B Streptococcus infection in a nonhuman primate model.Front Cell Infect Microbiol. 2024 Jan 4;13:1299644. doi: 10.3389/fcimb.2023.1299644. eCollection 2023. Front Cell Infect Microbiol. 2024. PMID: 38239507 Free PMC article.
-
Bacterial and Host Determinants of Group B Streptococcal Infection of the Neonate and Infant.Front Microbiol. 2022 Feb 21;13:820365. doi: 10.3389/fmicb.2022.820365. eCollection 2022. Front Microbiol. 2022. PMID: 35265059 Free PMC article. Review.
-
Function and contribution of two putative Enterococcus faecalis glycosaminoglycan degrading enzymes to bacteremia and catheter-associated urinary tract infection.Infect Immun. 2024 Jul 11;92(7):e0019924. doi: 10.1128/iai.00199-24. Epub 2024 Jun 6. Infect Immun. 2024. PMID: 38842305 Free PMC article.
-
Spatial transcriptome atlas reveals pulmonary microstructure-specific COVID-19 gene signatures in cynomolgus macaques.Commun Biol. 2023 Aug 28;6(1):879. doi: 10.1038/s42003-023-05253-8. Commun Biol. 2023. PMID: 37640792 Free PMC article.
-
Heterogeneity of the group B streptococcal type VII secretion system and influence on colonization of the female genital tract.bioRxiv [Preprint]. 2023 Jan 25:2023.01.25.525443. doi: 10.1101/2023.01.25.525443. bioRxiv. 2023. Update in: Mol Microbiol. 2023 Aug;120(2):258-275. doi: 10.1111/mmi.15115. PMID: 36747681 Free PMC article. Updated. Preprint.
References
-
- Institute of Medicine. 2007. In Behrman RE, Butler AS (ed), Preterm birth: causes, consequences, and prevention. Institute of Medicine, Washington, DC. doi:10.17226/11622.. - DOI
-
- Campbell JR, Hillier SL, Krohn MA, Ferrieri P, Zaleznik DF, Baker CJ. 2000. Group B streptococcal colonization and serotype-specific immunity in pregnant women at delivery. Obstet Gynecol 96:498–503. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
