

Adaptors as the regulators of HECT ubiquitin ligases
- PMID: 33402750
- PMCID: PMC7862271
- DOI: 10.1038/s41418-020-00707-6
Adaptors as the regulators of HECT ubiquitin ligases
Erratum in
-
Correction to: Adaptors as the regulators of HECT ubiquitin ligases.Cell Death Differ. 2021 Nov;28(11):3196. doi: 10.1038/s41418-021-00737-8. Cell Death Differ. 2021. PMID: 33483704 Free PMC article. No abstract available.
Abstract
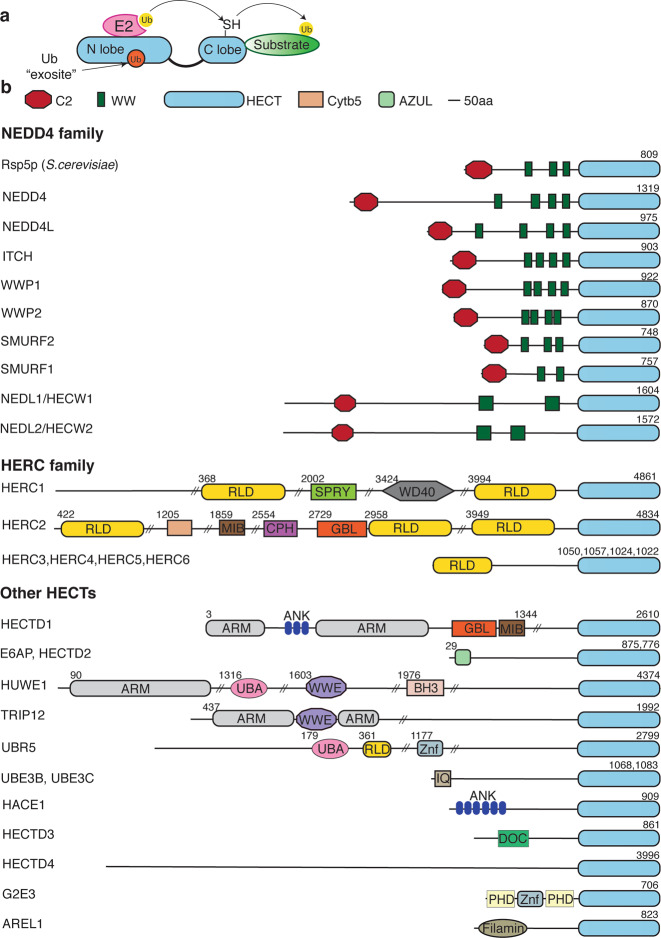
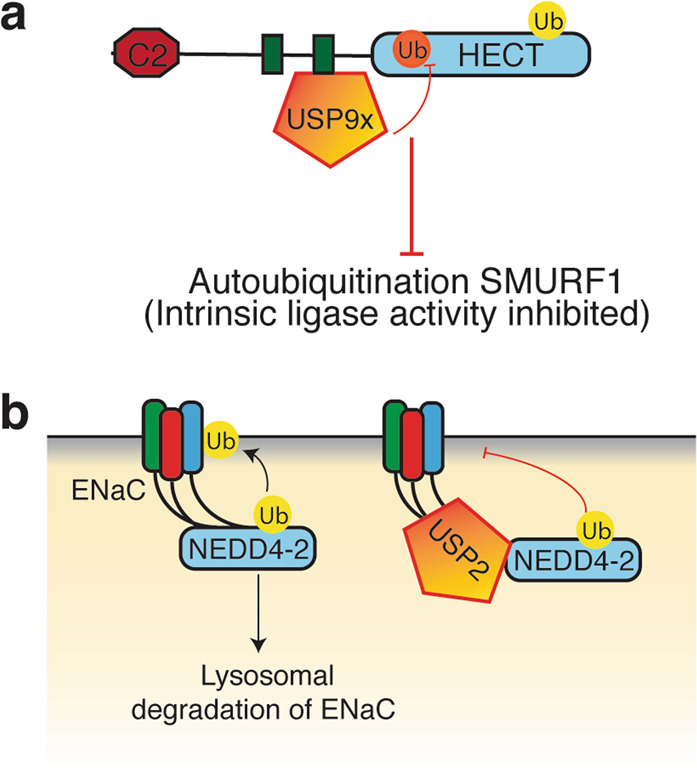
The HECT (homologous to E6AP C-terminus) ubiquitin ligases (E3s) are a small family of highly conserved enzymes involved in diverse cellular functions and pathological conditions. Characterised by a C-terminal HECT domain that accepts ubiquitin from E2 ubiquitin conjugating enzymes, these E3s regulate key signalling pathways. The activity and functional regulation of HECT E3s are controlled by several factors including post-translational modifications, inter- and intramolecular interactions and binding of co-activators and adaptor proteins. In this review, we focus on the regulation of HECT E3s by accessory proteins or adaptors and discuss various ways by which adaptors mediate their regulatory roles to affect physiological outcomes. We discuss common features that are conserved from yeast to mammals, regardless of the type of E3s as well as shed light on recent discoveries explaining some existing enigmas in the field.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures






References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
