

Dose-Dependent AGO1-Mediated Inhibition of the miRNA165/166 Pathway Modulates Stem Cell Maintenance in Arabidopsis Shoot Apical Meristem
- PMID: 33404539
- PMCID: PMC7747967
- DOI: 10.1016/j.xplc.2019.100002
Dose-Dependent AGO1-Mediated Inhibition of the miRNA165/166 Pathway Modulates Stem Cell Maintenance in Arabidopsis Shoot Apical Meristem
Abstract
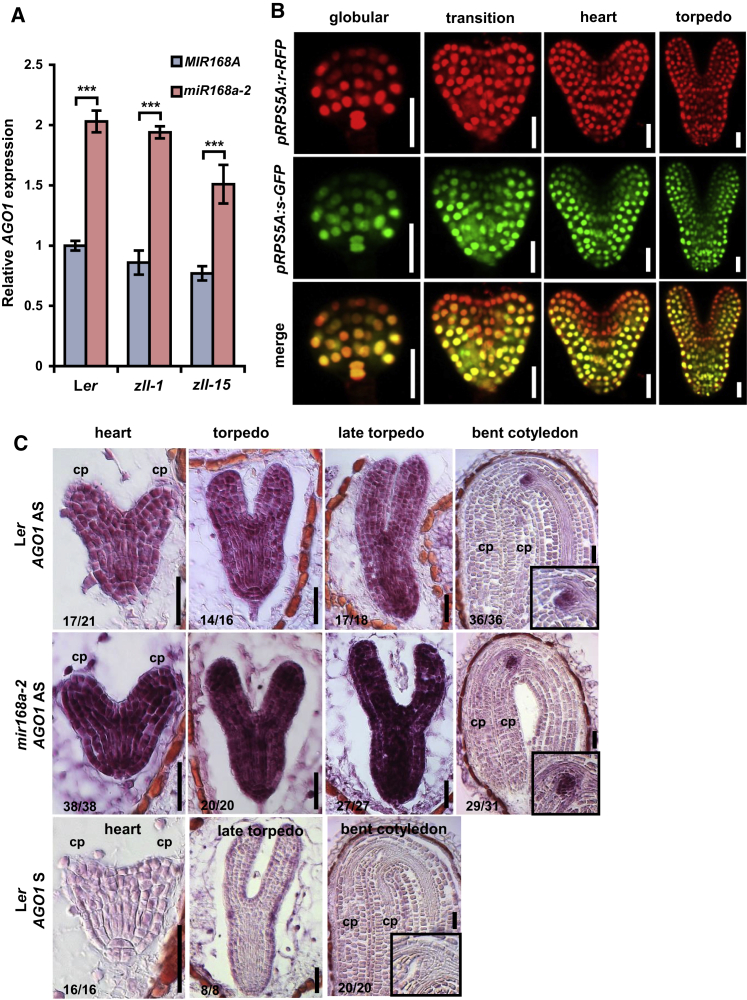
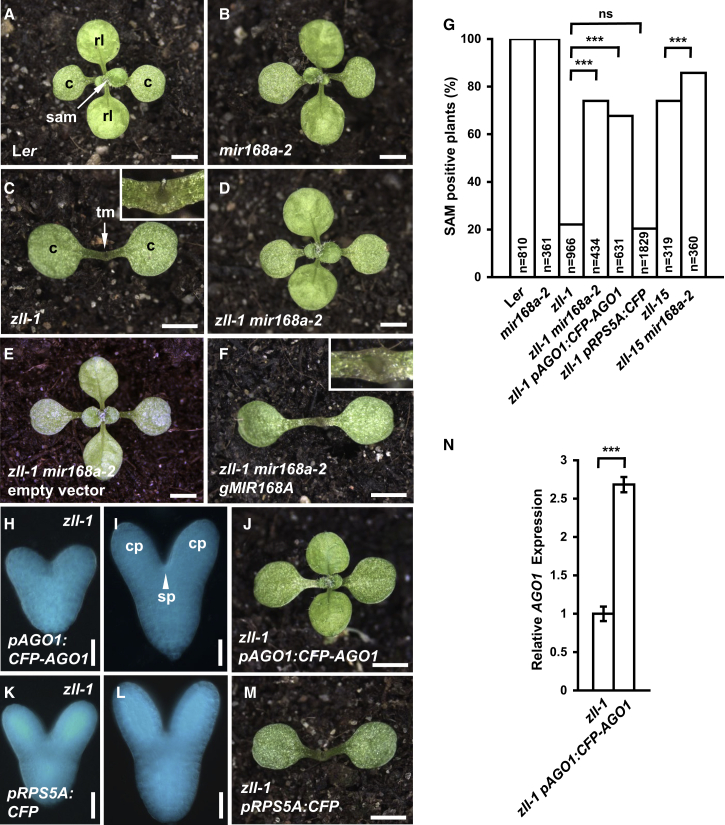
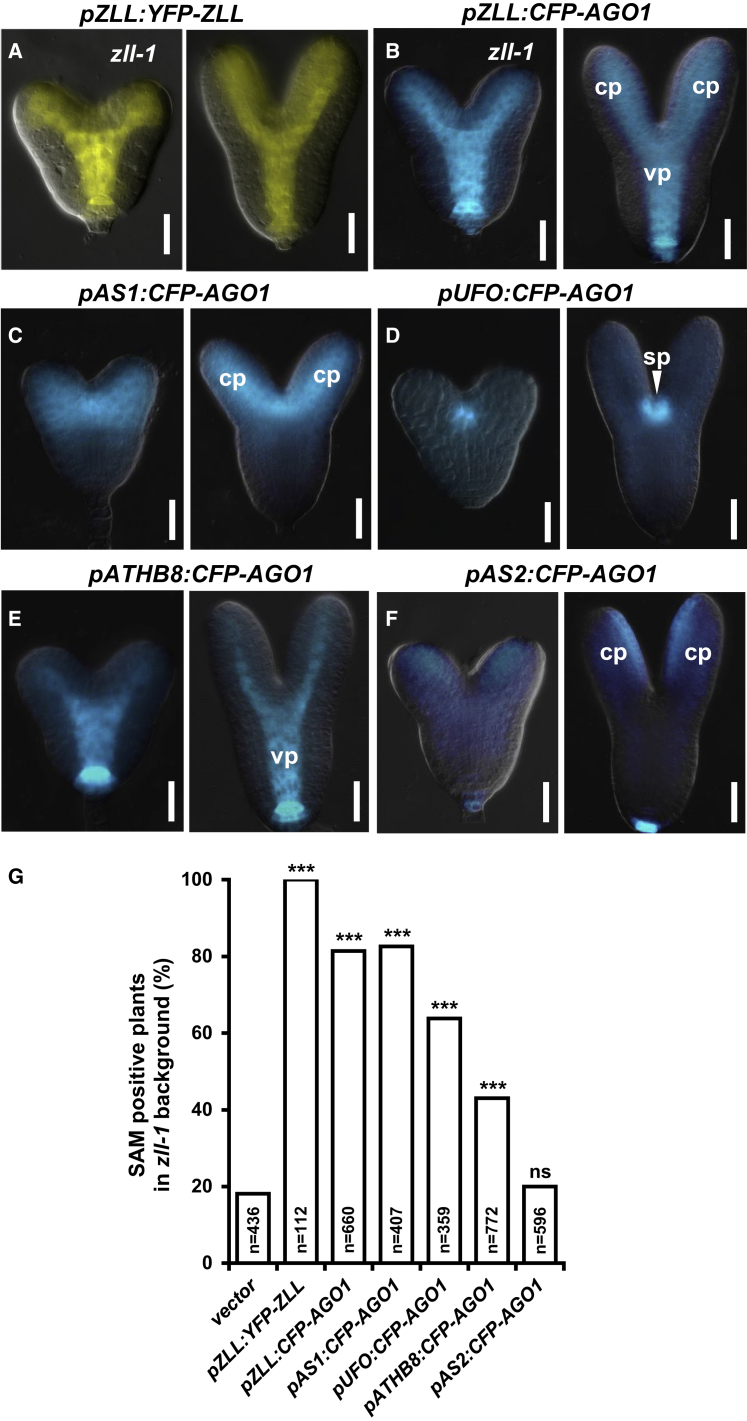
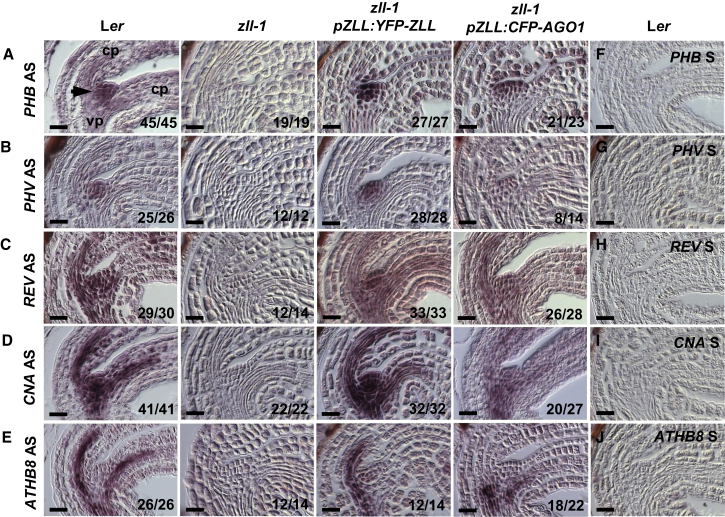
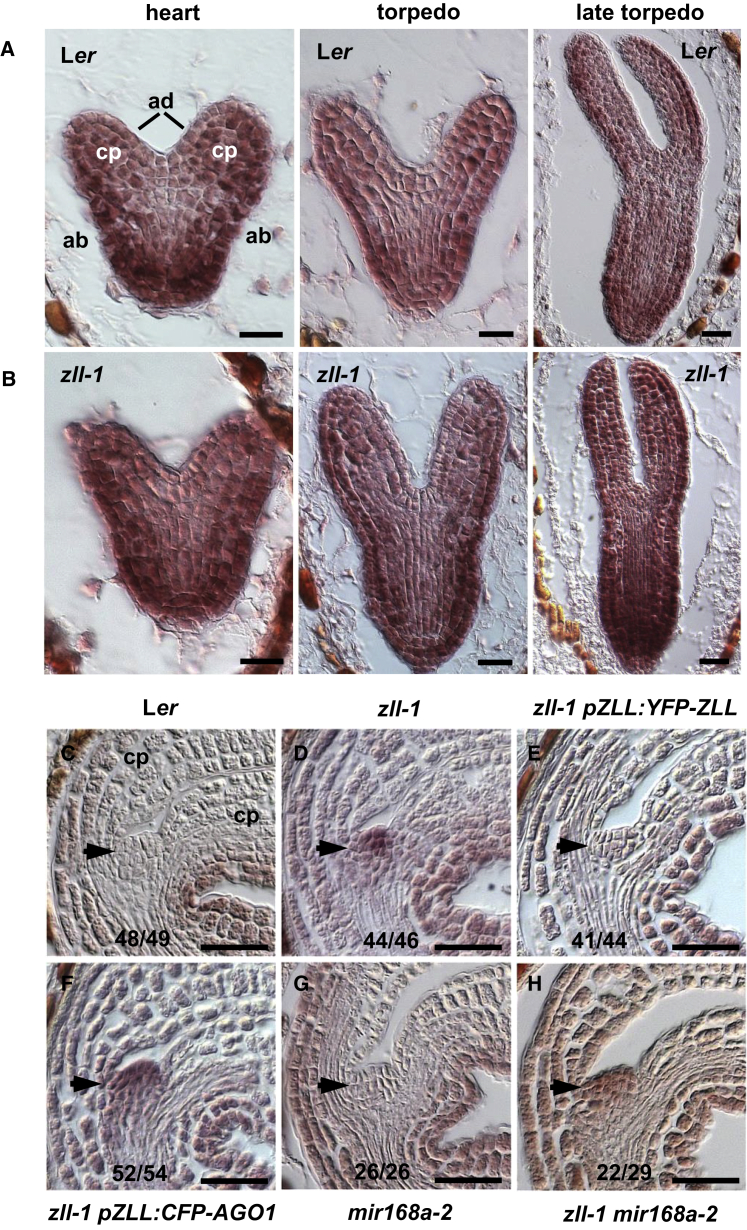
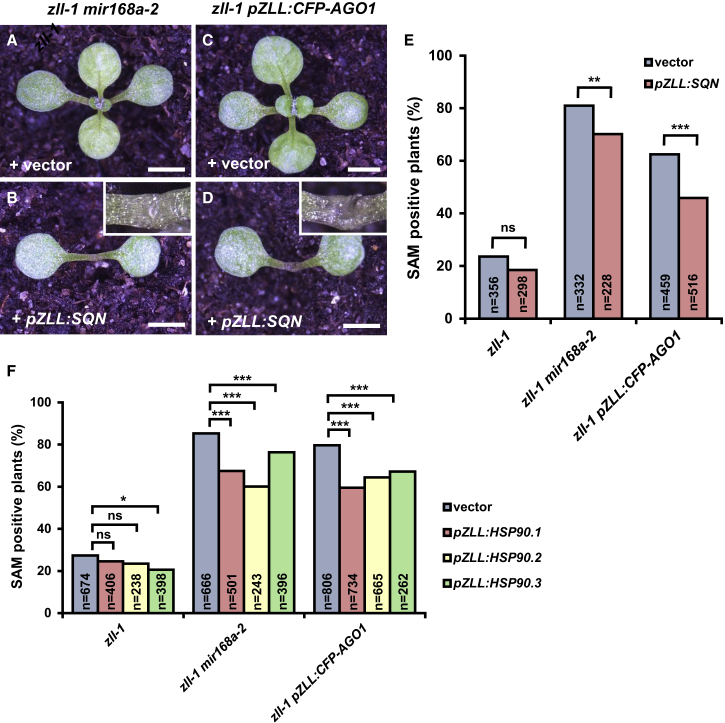
Pluripotent stem cells localized in proliferating growth centers, the meristems, are the origin of life-long organ formation and growth in higher plants. In the shoot apical meristem of Arabidopsis thaliana, the closely related ARGONAUTE proteins AGO1 and ZLL/AGO10 bind miR165/166 species to regulate mRNAs of HD-ZIP III transcription factors that are essential to maintaining stem cells. Several genetic studies showed that AGO1 and ZLL/AGO10 act redundantly to maintain stem cells. By contrast, the reported biochemical data suggested antagonistic functions: AGO1 utilizes miR165/166 to slice HD-ZIP III mRNAs, whereas ZLL/AGO10 promotes degradation of miR165/166 and thus stabilizes HD-ZIP III mRNAs. How these different functions are balanced in stem cell regulation has remained enigmatic. Here, we show that autorepression of AGO1 through miR168-mediated slicing of its own RNA is required to maintain the ability of AGO1 to suppress HD-ZIP III mRNAs. Increased AGO1 expression, either in the miR168a-2 mutant or by transgenic expression, inhibits this ability despite the presence of high levels of miR165/166, effectively uncoupling HD-ZIP III and miR165/166 expression. AGO1 activity can be restored, however, by increasing the levels of chaperones SQN and HSP90, which promote assembly of RNA-induced silencing complex (RISC). This suggests that cellular abundance of SQN and HSP chaperones limits AGO1-mediated RNA interference in shoot meristem stem cell regulation. Localized misexpression of AGO1 indicates that the cells surrounding the shoot meristem primordium play a crucial role in stem cell development. Taken together, our study provides a framework that reconciles biochemical and genetic data, showing that restriction of AGO1 levels by miR168-mediated autorepression is key to RISC homeostasis and the function of AGO1 in stem cell regulation.
Keywords: ARGONAUTE1; Arabidopsis; HD-ZIP III; ZWILLE (ARGONAUTE10); shoot meristem; stem cell.
© 2019 The Authors.
Figures
References
-
- Aichinger E., Kornet N., Friedrich T., Laux T. Plant stem cell niches. Annu. Rev. Plant Biol. 2012;63:615–636. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
