

Acetyl-CoA Synthetase 2: A Critical Linkage in Obesity-Induced Tumorigenesis in Myeloma
- PMID: 33406405
- PMCID: PMC7799390
- DOI: 10.1016/j.cmet.2020.12.011
Acetyl-CoA Synthetase 2: A Critical Linkage in Obesity-Induced Tumorigenesis in Myeloma
Abstract
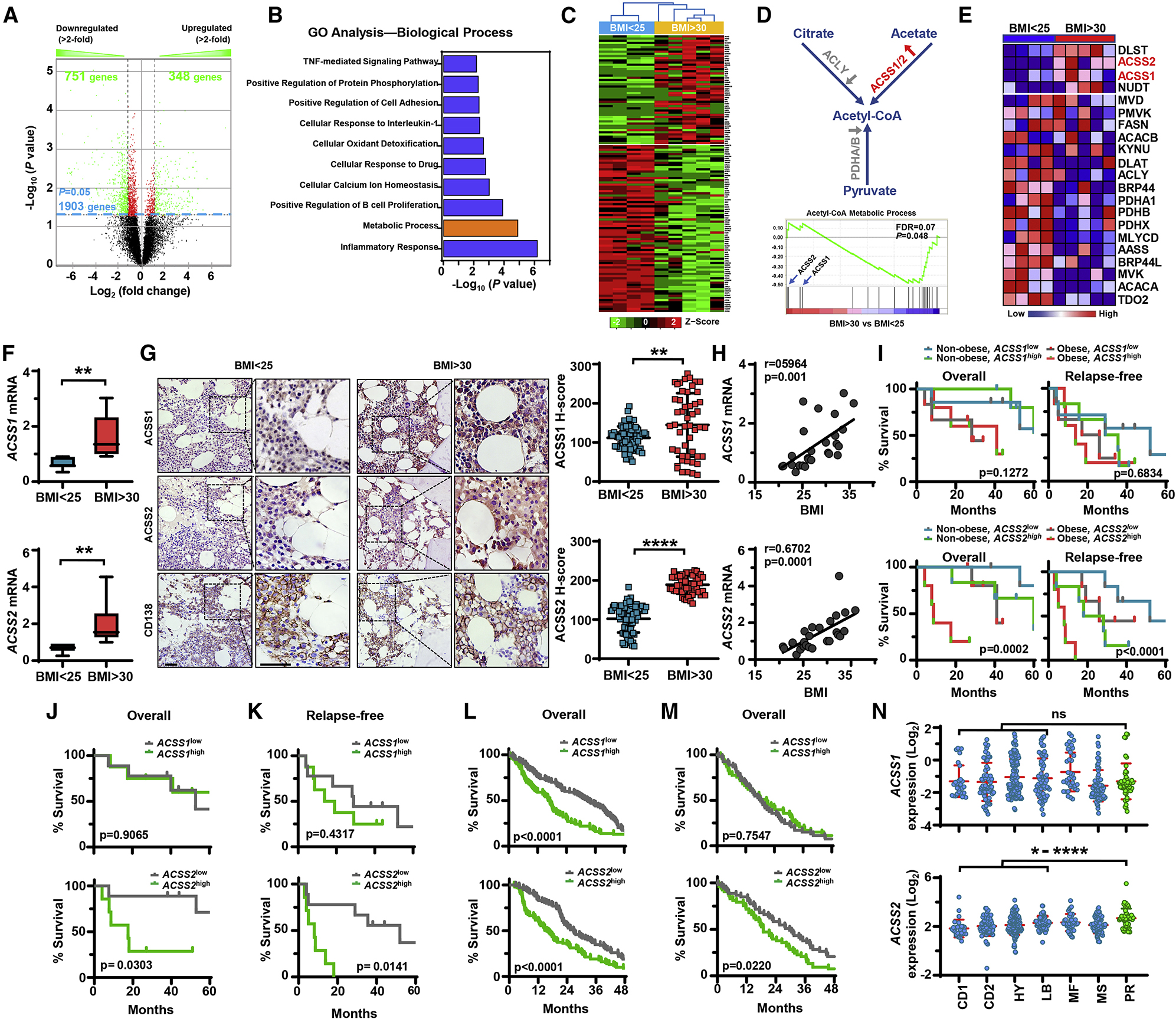
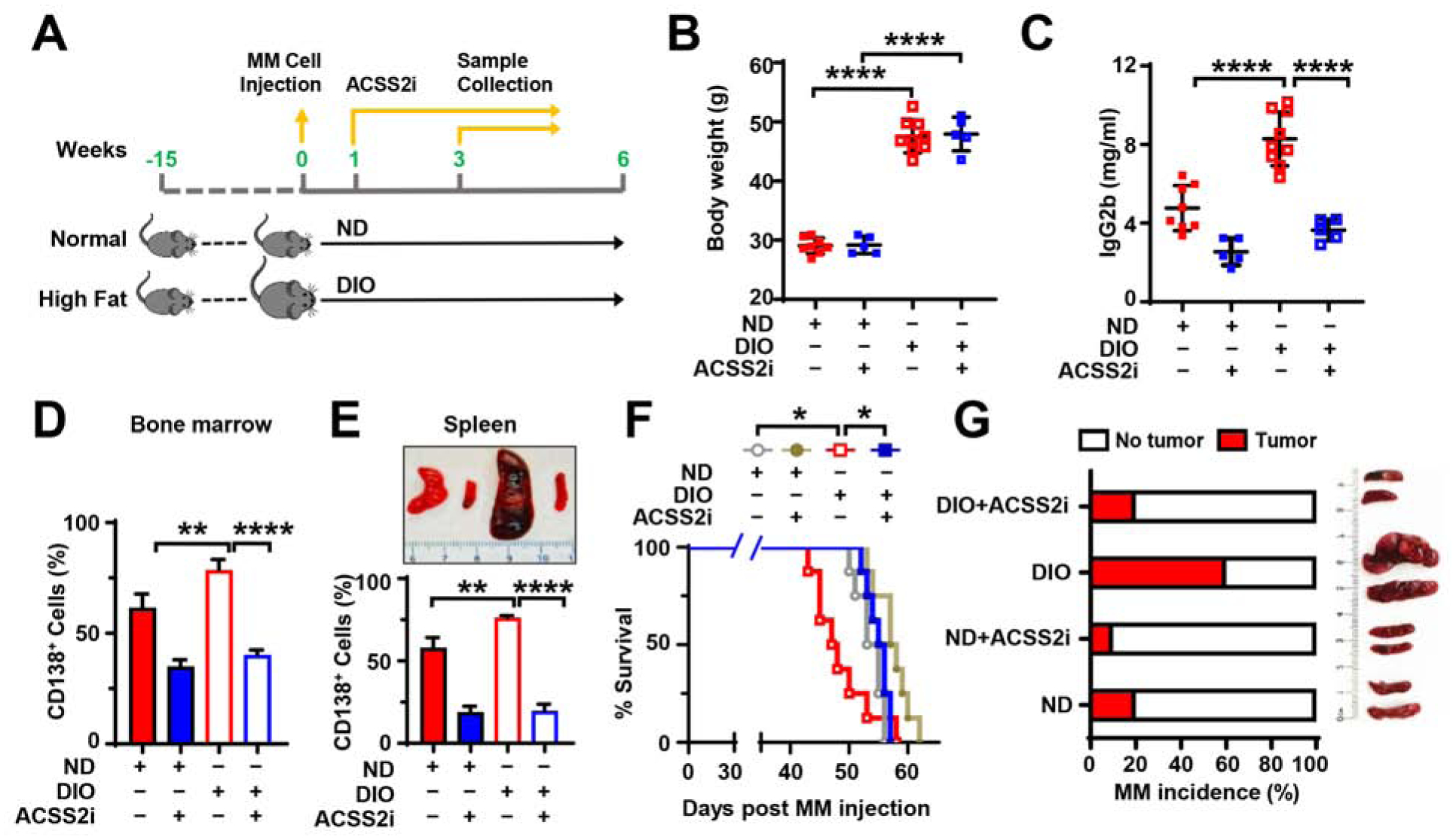
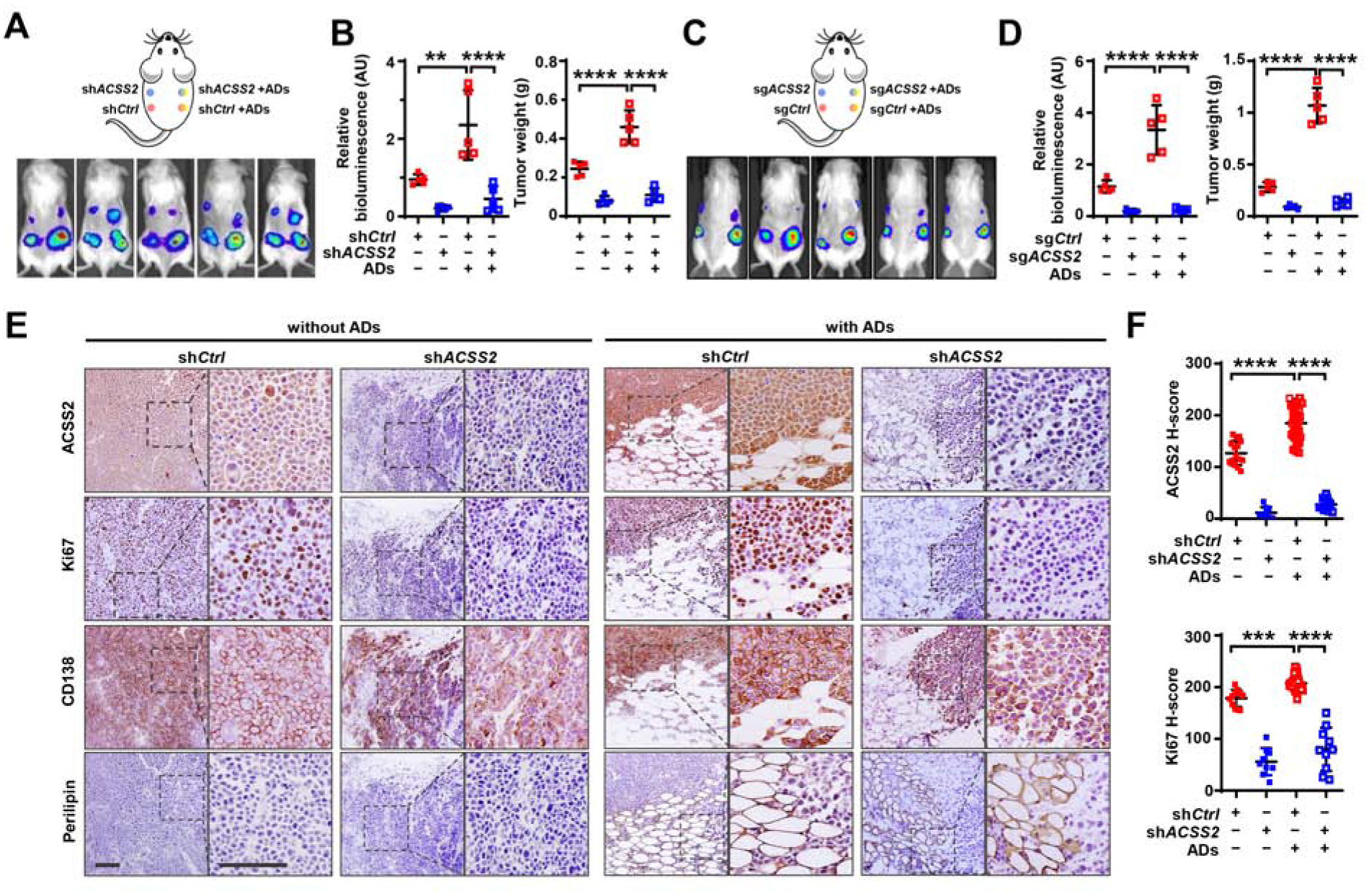
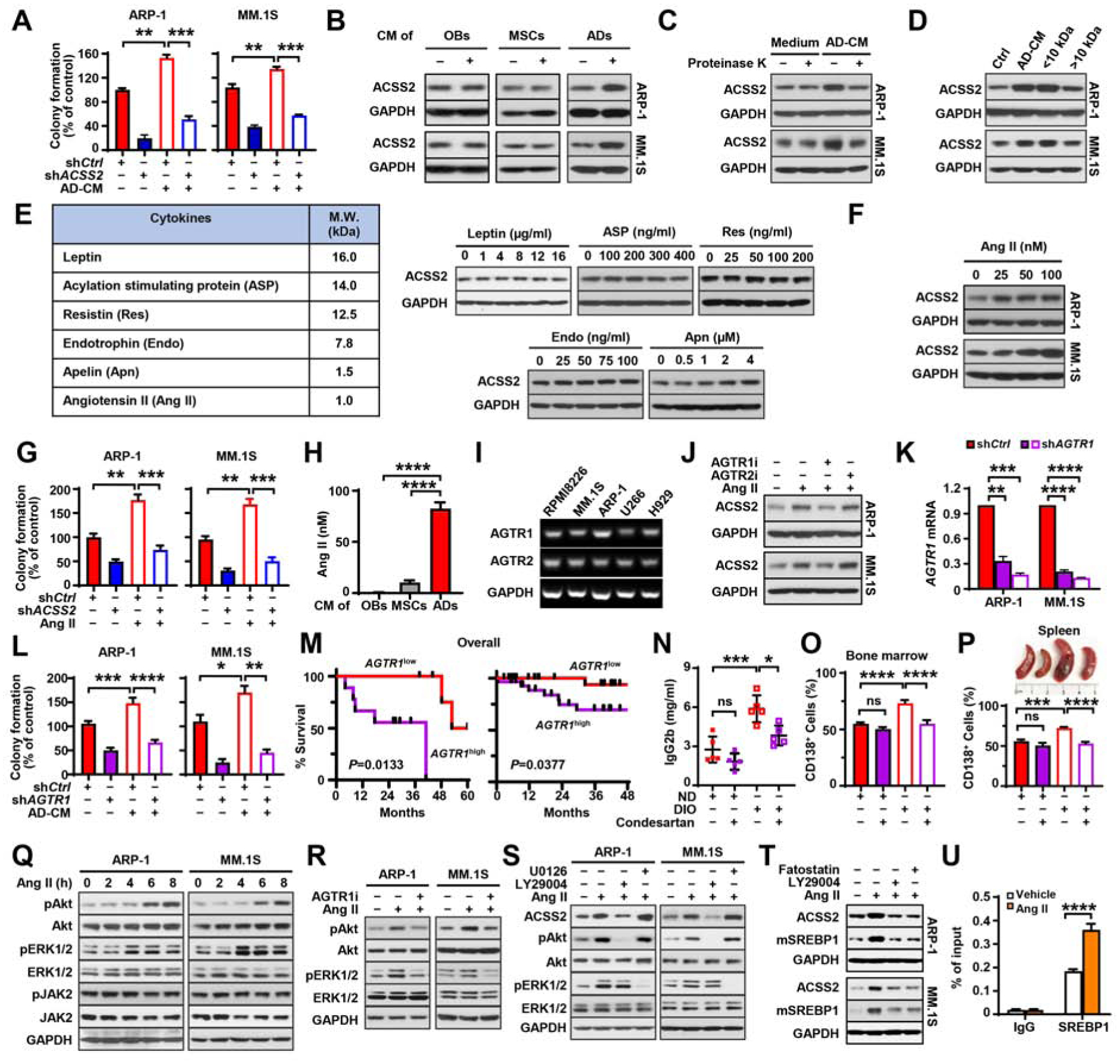
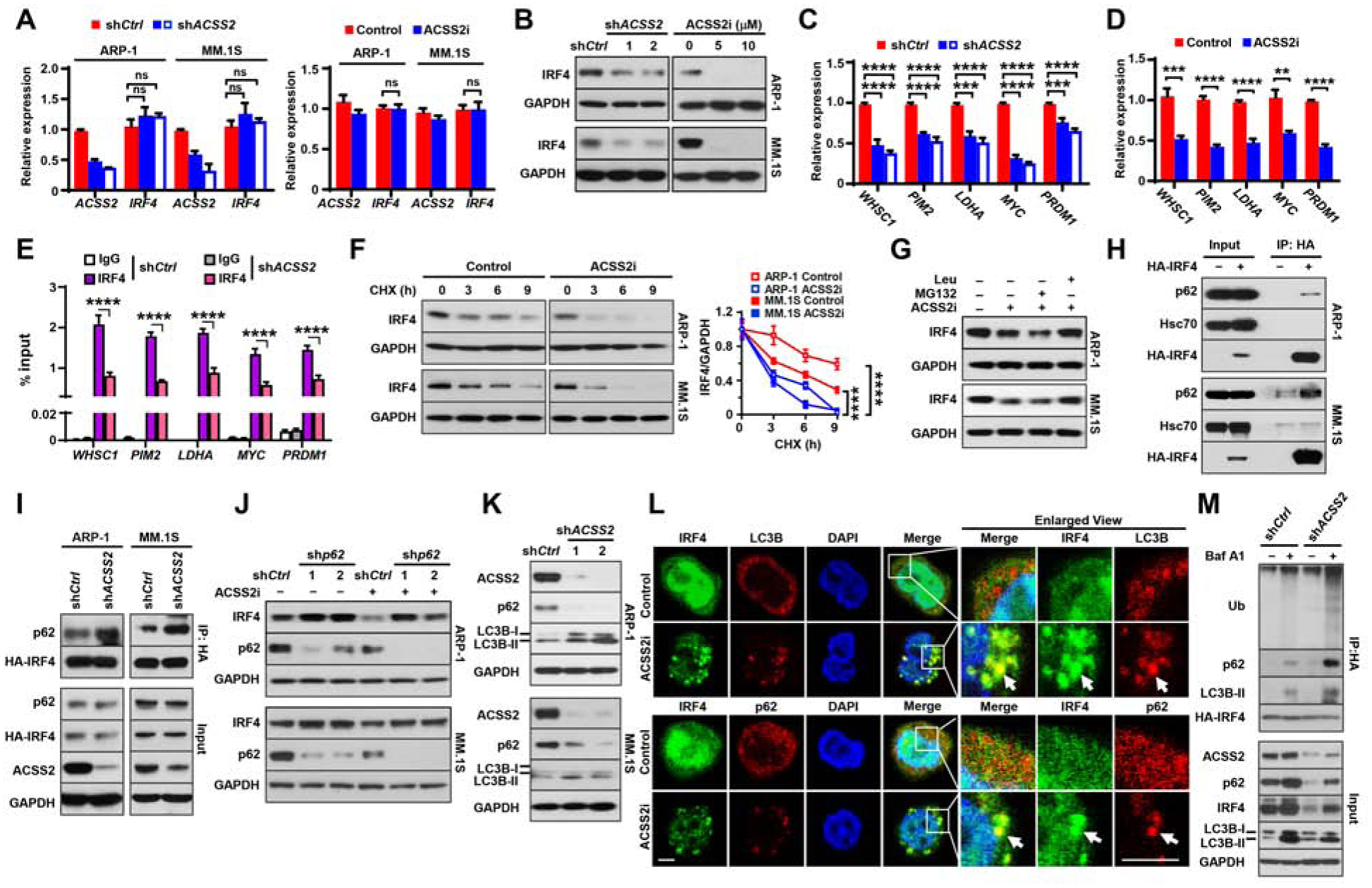
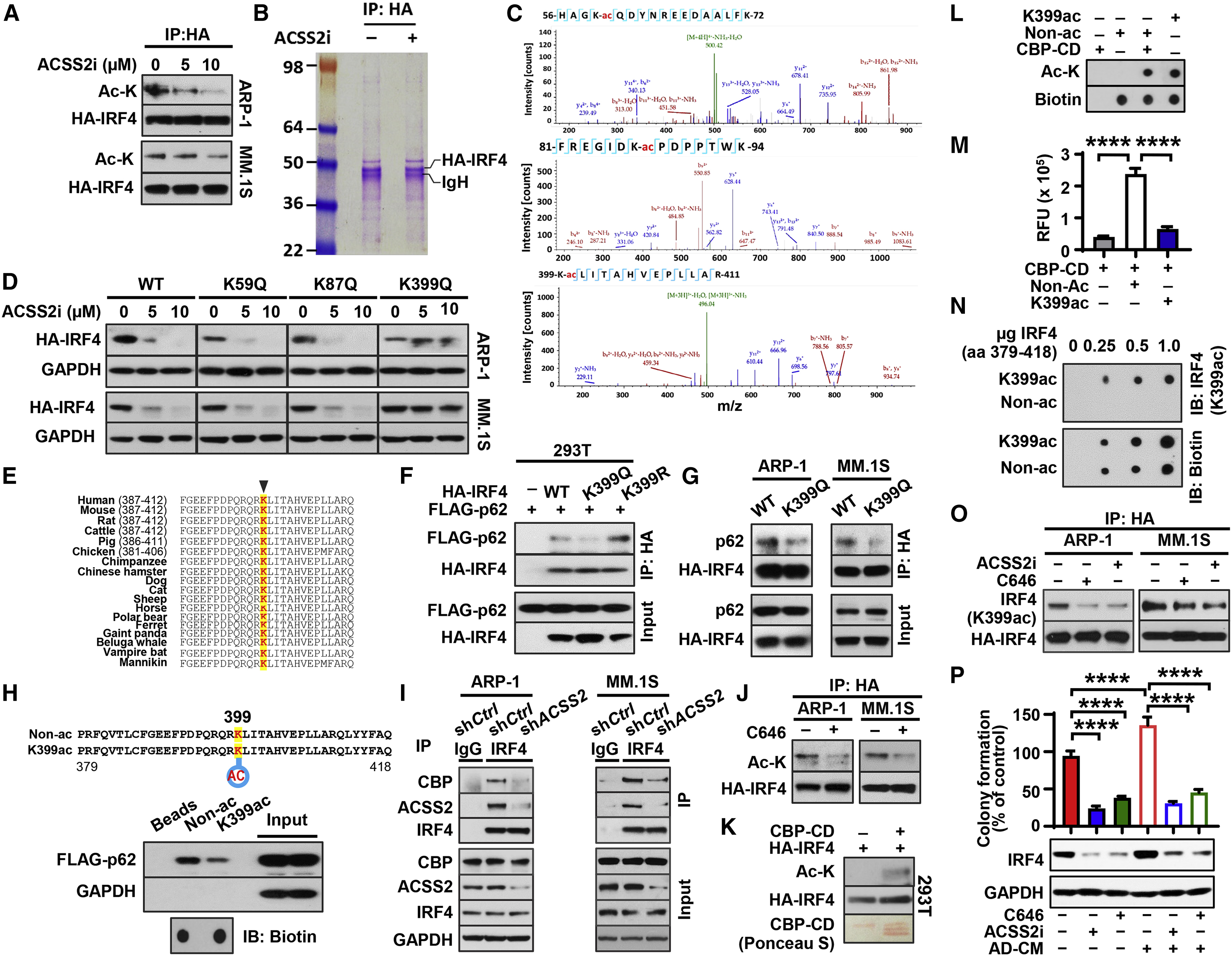
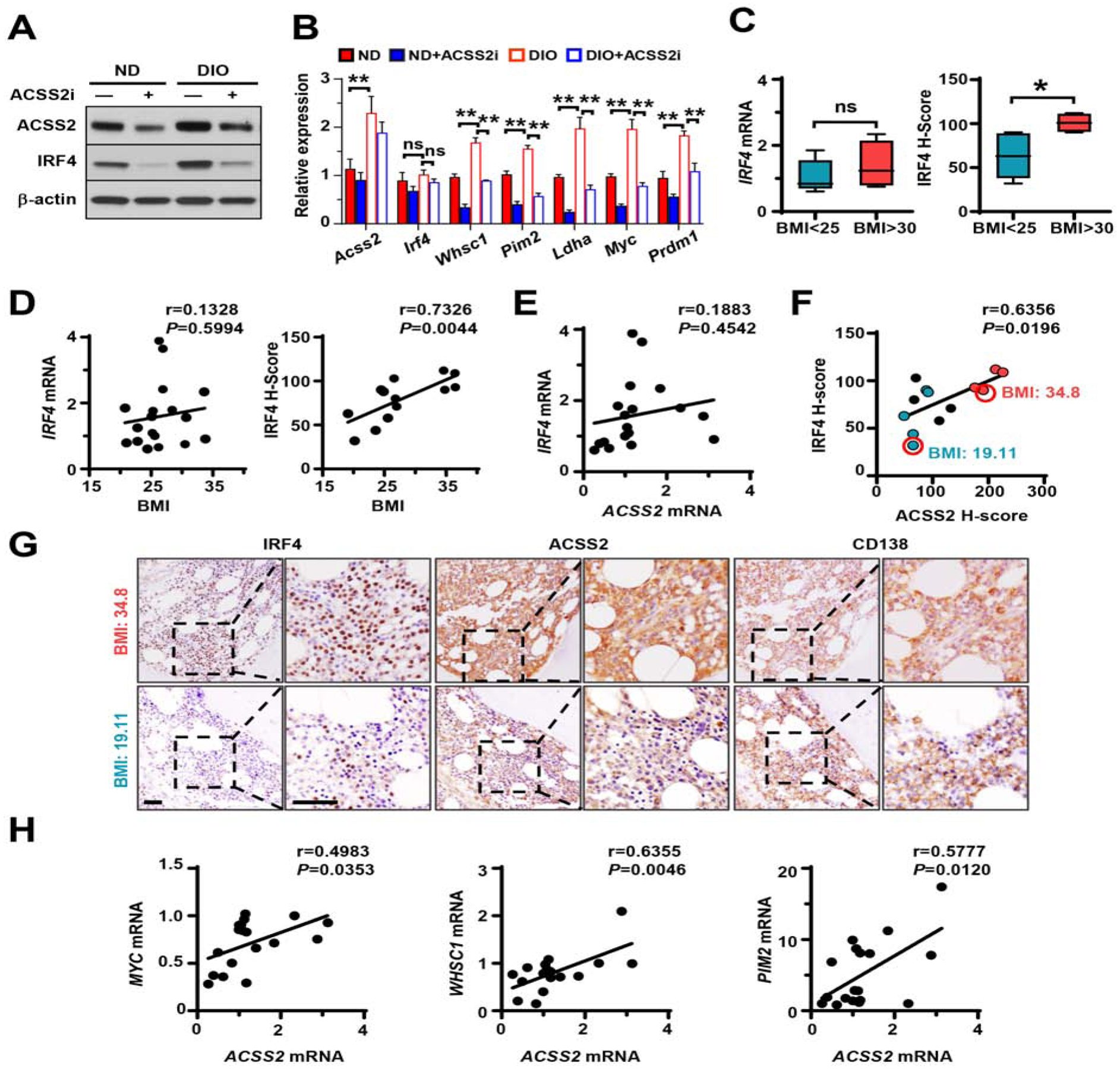
Obesity is often linked to malignancies including multiple myeloma, and the underlying mechanisms remain elusive. Here we showed that acetyl-CoA synthetase 2 (ACSS2) may be an important linker in obesity-related myeloma. ACSS2 is overexpressed in myeloma cells derived from obese patients and contributes to myeloma progression. We identified adipocyte-secreted angiotensin II as a direct cause of adiposity in increased ACSS2 expression. ACSS2 interacts with oncoprotein interferon regulatory factor 4 (IRF4), and enhances IRF4 stability and IRF4-mediated gene transcription through activation of acetylation. The importance of ACSS2 overexpression in myeloma is confirmed by the finding that an inhibitor of ACSS2 reduces myeloma growth both in vitro and in a diet-induced obese mouse model. Our findings demonstrate a key impact for obesity-induced ACSS2 on the progression of myeloma. Given the central role of ACSS2 in many tumors, this mechanism could be important to other obesity-related malignancies.
Keywords: ACSS2; IRF4; adipocytes; angiotensin II; autophagy; lysine acetylation; multiple myeloma; obesity.
Copyright © 2020 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of Interests The authors declare no conflicts of interest.
Figures
References
-
- Azizi M, and Menard J (2004). Combined blockade of the renin-angiotensin system with angiotensin-converting enzyme inhibitors and angiotensin II type 1 receptor antagonists. Circulation 109, 2492–2499. - PubMed
-
- Bianchini F, Kaaks R, and Vainio H (2002). Overweight, obesity, and cancer risk. Lancet Oncology 3, 565–574. - PubMed
-
- Billet S, Aguilar F, Baudry C, and Clauser E (2008). Role of angiotensin II AT1 receptor activation in cardiovascular diseases. Kidney Int 74, 1379–1384. - PubMed
-
- Blair CK, Cerhan JR, Folsom AR, and Ross JA (2005). Anthropometric characteristics and risk of multiple myeloma. Epidemiology 16, 691–694. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
