

Genome-wide occupancy reveals the localization of H1T2 (H1fnt) to repeat regions and a subset of transcriptionally active chromatin domains in rat spermatids
- PMID: 33407810
- PMCID: PMC7788777
- DOI: 10.1186/s13072-020-00376-2
Genome-wide occupancy reveals the localization of H1T2 (H1fnt) to repeat regions and a subset of transcriptionally active chromatin domains in rat spermatids
Abstract
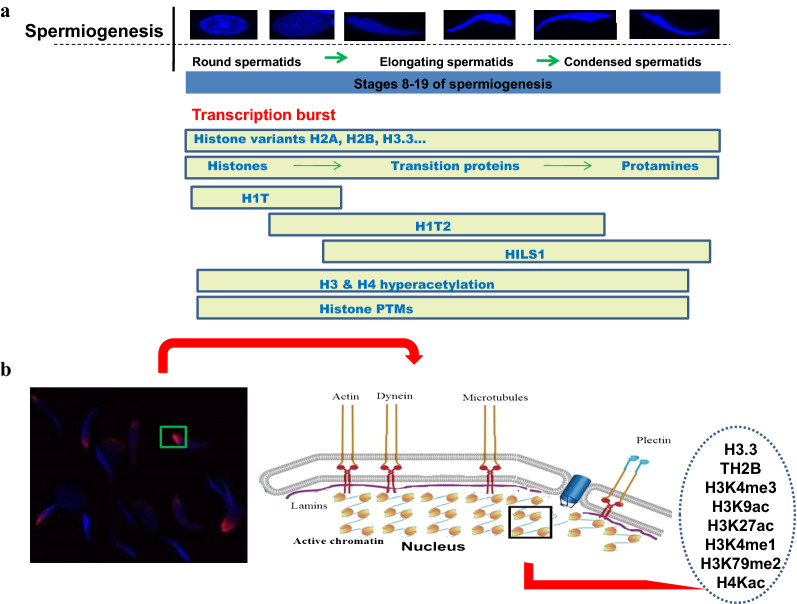
Background: H1T2/H1FNT is a germ cell-specific linker histone variant expressed during spermiogenesis specifically in round and elongating spermatids. Infertile phenotype of homozygous H1T2 mutant male mice revealed the essential function of H1T2 for the DNA condensation and histone-to-protamine replacement in spermiogenesis. However, the mechanism by which H1T2 imparts the inherent polarity within spermatid nucleus including the additional protein partners and the genomic domains occupied by this linker histone are unknown.
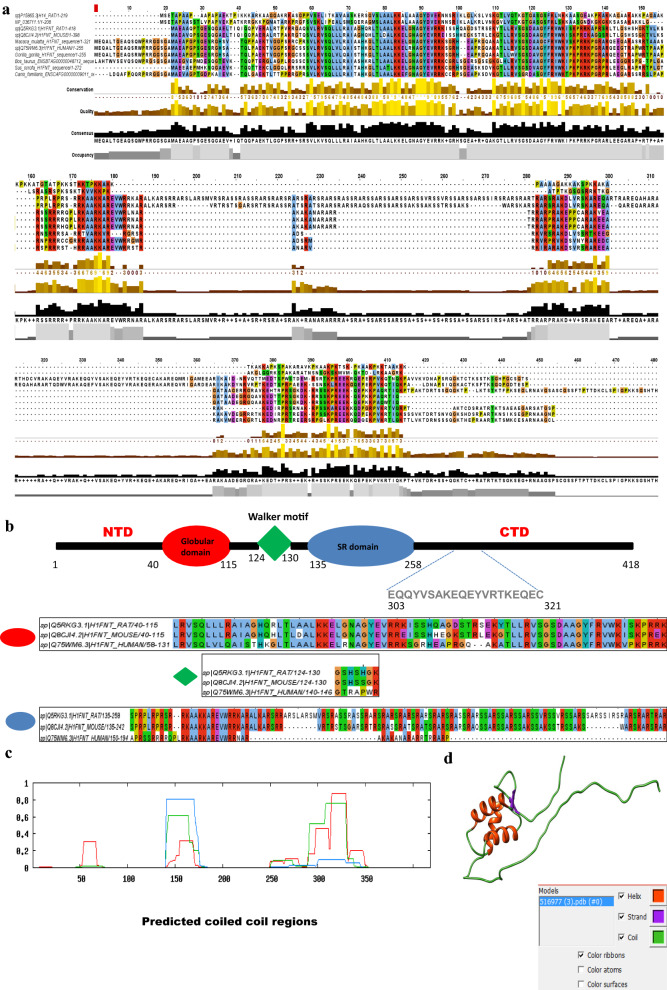
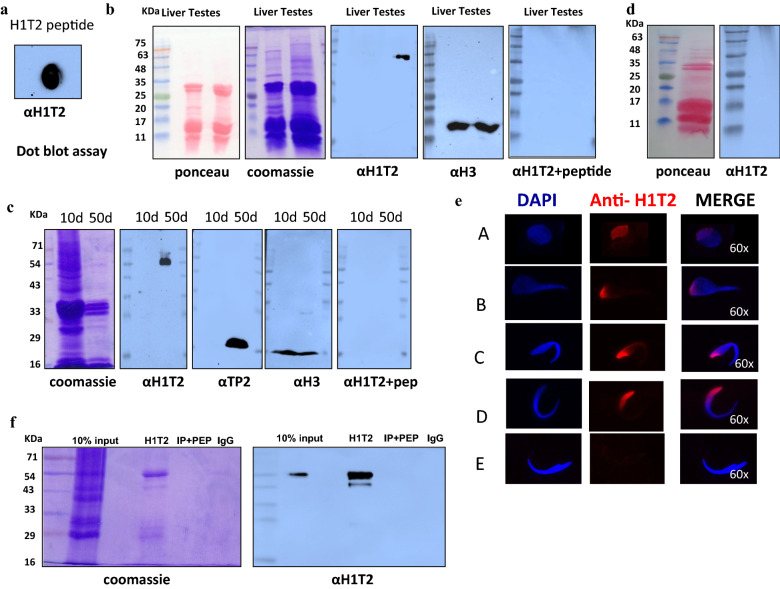
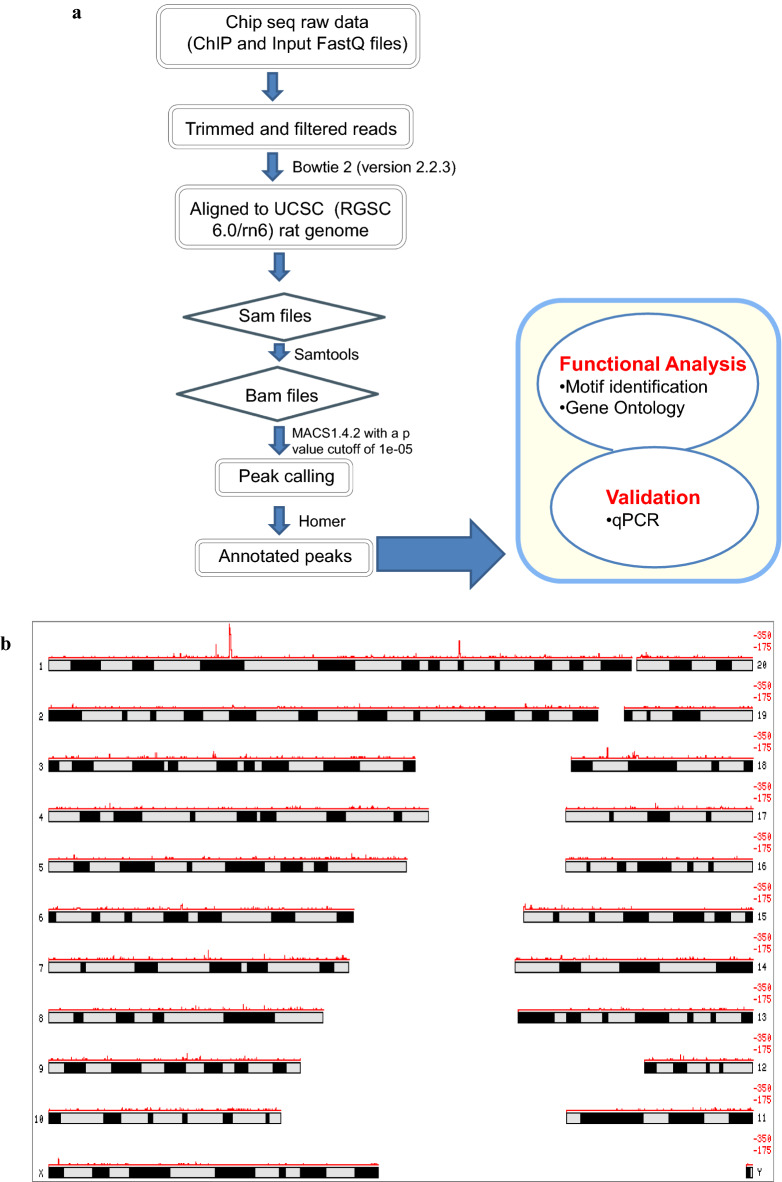
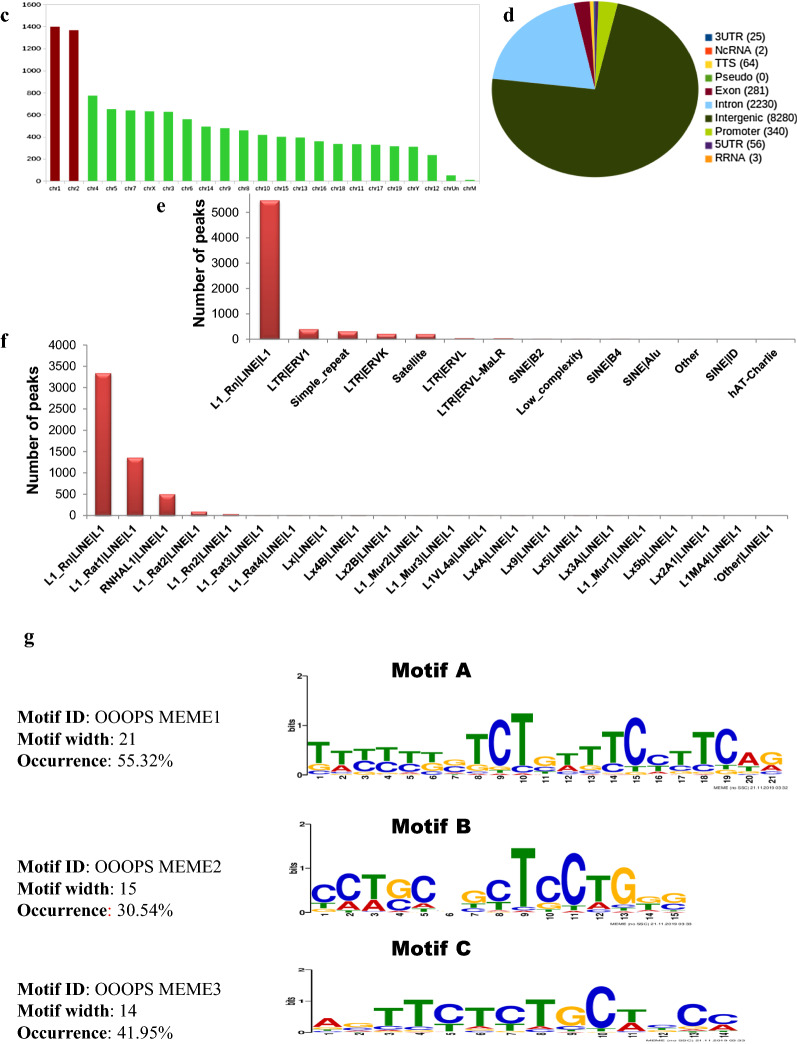
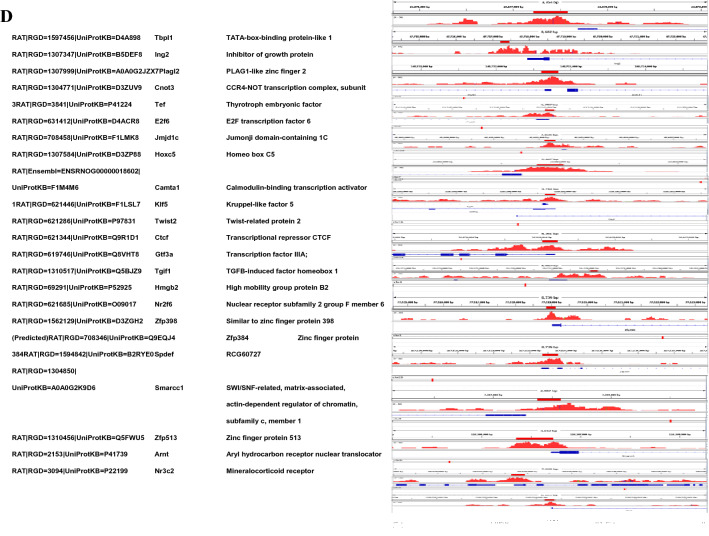
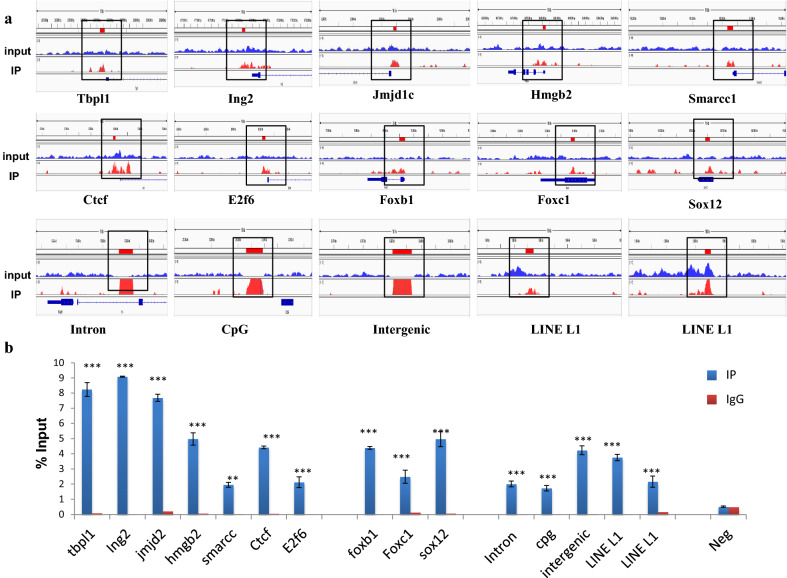
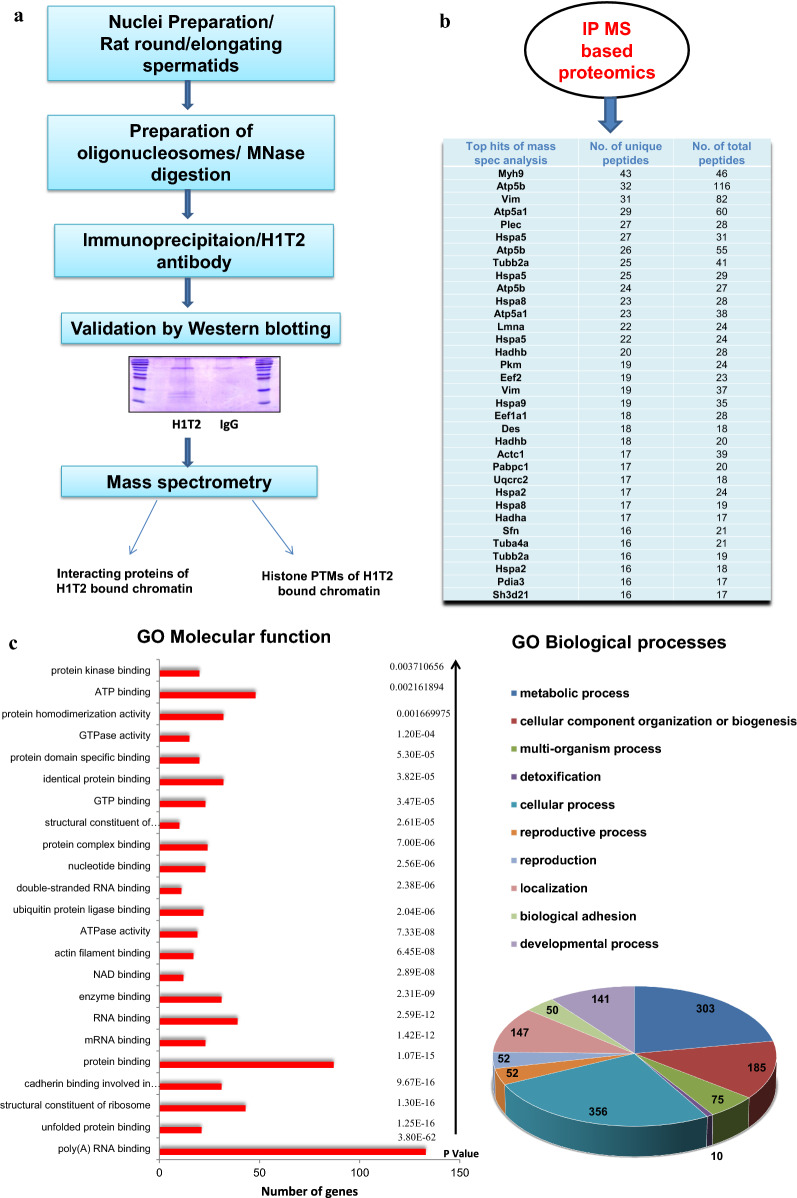
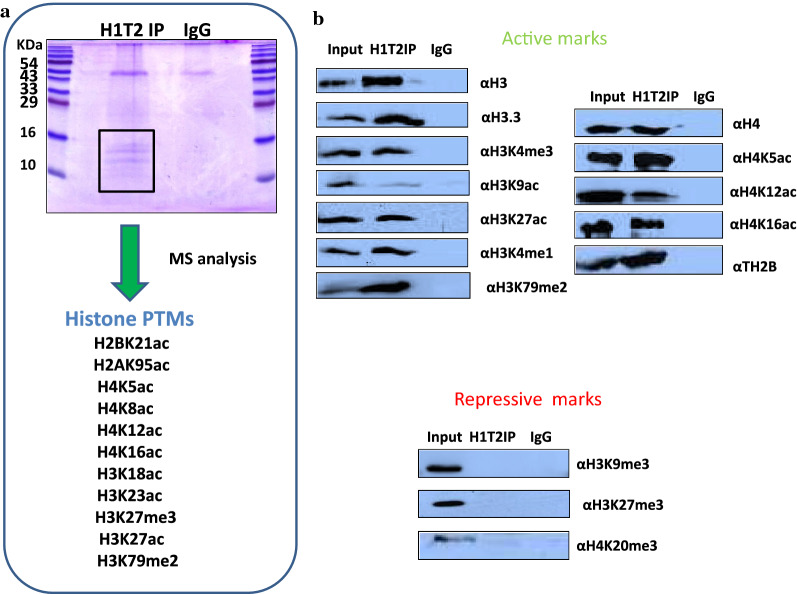
Results: Sequence analysis revealed the presence of Walker motif, SR domains and putative coiled-coil domains in the C-terminal domain of rat H1T2 protein. Genome-wide occupancy analysis using highly specific antibody against the CTD of H1T2 demonstrated the binding of H1T2 to the LINE L1 repeat elements and to a significant percentage of the genic regions (promoter-TSS, exons and introns) of the rat spermatid genome. Immunoprecipitation followed by mass spectrometry analysis revealed the open chromatin architecture of H1T2 occupied chromatin encompassing the H4 acetylation and other histone PTMs characteristic of transcriptionally active chromatin. In addition, the present study has identified the interacting protein partners of H1T2-associated chromatin mainly as nucleo-skeleton components, RNA-binding proteins and chaperones.
Conclusions: Linker histone H1T2 possesses unique domain architecture which can account for the specific functions associated with chromatin remodeling events facilitating the initiation of histone to transition proteins/protamine transition in the polar apical spermatid genome. Our results directly establish the unique function of H1T2 in nuclear shaping associated with spermiogenesis by mediating the interaction between chromatin and nucleo-skeleton, positioning the epigenetically specialized chromatin domains involved in transcription coupled histone replacement initiation towards the apical pole of round/elongating spermatids.
Keywords: ChIP-sequencing; Histone PTMs; Linker histone; Spermatid; Spermiogenesis.
Conflict of interest statement
The authors declare that they have no competing interests with the contents of this article.
Figures


Similar articles
-
Polar nuclear localization of H1T2, a histone H1 variant, required for spermatid elongation and DNA condensation during spermiogenesis.Proc Natl Acad Sci U S A. 2005 Feb 22;102(8):2808-13. doi: 10.1073/pnas.0406060102. Epub 2005 Feb 14. Proc Natl Acad Sci U S A. 2005. PMID: 15710904 Free PMC article.
-
Changes in intranuclear chromatin architecture induce bipolar nuclear localization of histone variant H1T2 in male haploid spermatids.Dev Biol. 2006 Aug 1;296(1):231-238. doi: 10.1016/j.ydbio.2006.04.458. Dev Biol. 2006. PMID: 16765935
-
Spermatid-specific linker histone HILS1 is a poor condenser of DNA and chromatin and preferentially associates with LINE-1 elements.Epigenetics Chromatin. 2018 Aug 1;11(1):43. doi: 10.1186/s13072-018-0214-0. Epigenetics Chromatin. 2018. PMID: 30068355 Free PMC article.
-
Sperm chromatin protamination: an endocrine perspective.Protein Pept Lett. 2011 Aug;18(8):786-801. doi: 10.2174/092986611795714005. Protein Pept Lett. 2011. PMID: 21443490 Review.
-
Chromatin dynamics during spermiogenesis.Biochim Biophys Acta. 2014 Mar;1839(3):155-68. doi: 10.1016/j.bbagrm.2013.08.004. Epub 2013 Sep 30. Biochim Biophys Acta. 2014. PMID: 24091090 Review.
Cited by
-
A-MYB and BRDT-dependent RNA Polymerase II pause release orchestrates transcriptional regulation in mammalian meiosis.Nat Commun. 2023 Mar 29;14(1):1753. doi: 10.1038/s41467-023-37408-w. Nat Commun. 2023. PMID: 36990976 Free PMC article.
-
Heterogeneity in the Epigenetic Landscape of Murine Testis-Specific Histone Variants TH2A and TH2B Sharing the Same Bi-Directional Promoter.Front Cell Dev Biol. 2021 Dec 6;9:755751. doi: 10.3389/fcell.2021.755751. eCollection 2021. Front Cell Dev Biol. 2021. PMID: 34938732 Free PMC article.
-
Casting histone variants during mammalian reproduction.Chromosoma. 2023 Sep;132(3):153-165. doi: 10.1007/s00412-023-00803-9. Epub 2023 Jun 22. Chromosoma. 2023. PMID: 37347315 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
