

Roles of Cholesterol in Early and Late Steps of the Nipah Virus Membrane Fusion Cascade
- PMID: 33408170
- PMCID: PMC8094960
- DOI: 10.1128/JVI.02323-20
Roles of Cholesterol in Early and Late Steps of the Nipah Virus Membrane Fusion Cascade
Abstract
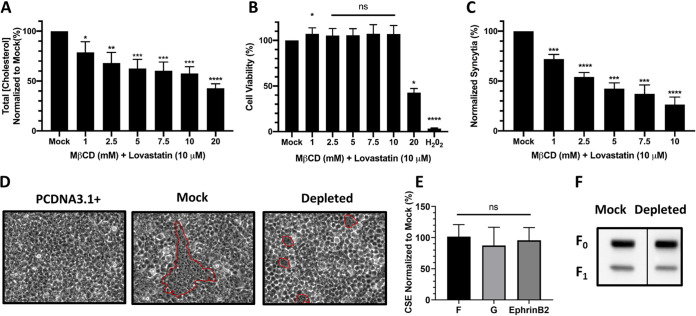
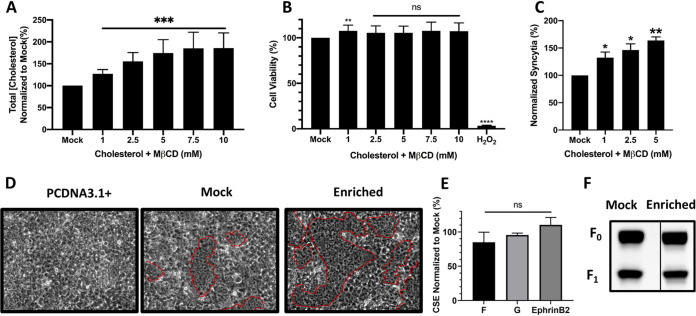
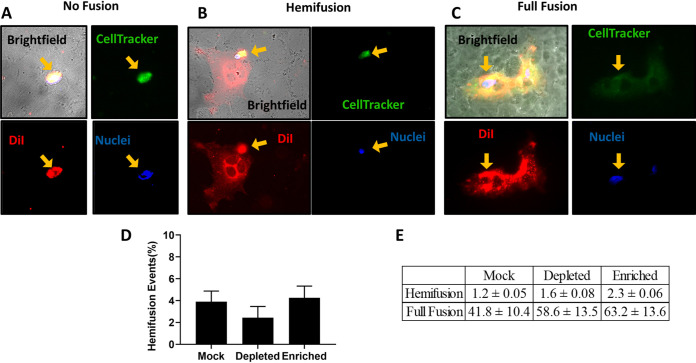
Cholesterol has been implicated in various viral life cycle steps for different enveloped viruses, including viral entry into host cells, cell-cell fusion, and viral budding from infected cells. Enveloped viruses acquire their membranes from their host cells. Although cholesterol has been associated with the binding and entry of various enveloped viruses into cells, cholesterol's exact function in the viral-cell membrane fusion process remains largely elusive, particularly for the paramyxoviruses. Furthermore, paramyxoviral fusion occurs at the host cell membrane and is essential for both virus entry (virus-cell fusion) and syncytium formation (cell-cell fusion), central to viral pathogenicity. Nipah virus (NiV) is a deadly member of the Paramyxoviridae family, which also includes Hendra, measles, mumps, human parainfluenza, and various veterinary viruses. The zoonotic NiV causes severe encephalitis, vasculopathy, and respiratory symptoms, leading to a high mortality rate in humans. We used NiV as a model to study the role of membrane cholesterol in paramyxoviral membrane fusion. We used a combination of methyl-beta cyclodextrin (MβCD), lovastatin, and cholesterol to deplete or enrich cell membrane cholesterol outside cytotoxic concentrations. We found that the levels of cellular membrane cholesterol directly correlated with the levels of cell-cell fusion induced. These phenotypes were paralleled using NiV/vesicular stomatitis virus (VSV)-pseudotyped viral infection assays. Remarkably, our mechanistic studies revealed that cholesterol reduces an early F-triggering step but enhances a late fusion pore formation step in the NiV membrane fusion cascade. Thus, our results expand our mechanistic understanding of the paramyxoviral/henipaviral entry and cell-cell fusion processes.IMPORTANCE Cholesterol has been implicated in various steps of the viral life cycle for different enveloped viruses. Nipah virus (NiV) is a highly pathogenic enveloped virus in the Henipavirus genus within the Paramyxoviridae family, capable of causing a high mortality rate in humans and high morbidity in domestic and agriculturally important animals. The role of cholesterol for NiV or the henipaviruses is unknown. Here, we show that the levels of cholesterol influence the levels of NiV-induced cell-cell membrane fusion during syncytium formation and virus-cell membrane fusion during viral entry. Furthermore, the specific role of cholesterol in membrane fusion is not well defined for the paramyxoviruses. We show that the levels of cholesterol affect an early F-triggering step and a late fusion pore formation step during the membrane fusion cascade. Thus, our results expand our mechanistic understanding of the viral entry and cell-cell fusion processes, which may aid the development of antivirals.
Keywords: Nipah virus; cholesterol; entry; fusion; henipavirus; membrane; paramyxovirus.
Copyright © 2021 American Society for Microbiology.
Figures
Similar articles
-
Third Helical Domain of the Nipah Virus Fusion Glycoprotein Modulates both Early and Late Steps in the Membrane Fusion Cascade.J Virol. 2020 Sep 15;94(19):e00644-20. doi: 10.1128/JVI.00644-20. Print 2020 Sep 15. J Virol. 2020. PMID: 32669342 Free PMC article.
-
Novel Roles of the N1 Loop and N4 Alpha-Helical Region of the Nipah Virus Fusion Glycoprotein in Modulating Early and Late Steps of the Membrane Fusion Cascade.J Virol. 2021 Apr 12;95(9):e01707-20. doi: 10.1128/JVI.01707-20. Print 2021 Apr 12. J Virol. 2021. PMID: 33568505 Free PMC article.
-
Novel Roles of the Nipah Virus Attachment Glycoprotein and Its Mobility in Early and Late Membrane Fusion Steps.mBio. 2022 Jun 28;13(3):e0322221. doi: 10.1128/mbio.03222-21. Epub 2022 May 4. mBio. 2022. PMID: 35506666 Free PMC article.
-
Pseudotyped Virus for Henipavirus.Adv Exp Med Biol. 2023;1407:175-190. doi: 10.1007/978-981-99-0113-5_9. Adv Exp Med Biol. 2023. PMID: 36920697 Review.
-
Henipavirus mediated membrane fusion, virus entry and targeted therapeutics.Viruses. 2012 Feb;4(2):280-308. doi: 10.3390/v4020280. Epub 2012 Feb 13. Viruses. 2012. PMID: 22470837 Free PMC article. Review.
Cited by
-
Interaction of Nipah Virus F and G with the Cellular Protein Cortactin Discovered by a Proximity Interactome Assay.Int J Mol Sci. 2024 Apr 8;25(7):4112. doi: 10.3390/ijms25074112. Int J Mol Sci. 2024. PMID: 38612921 Free PMC article.
-
Development of Fusion-Based Assay as a Drug Screening Platform for Nipah Virus Utilizing Baculovirus Expression Vector System.Int J Mol Sci. 2024 Aug 22;25(16):9102. doi: 10.3390/ijms25169102. Int J Mol Sci. 2024. PMID: 39201788 Free PMC article.
-
Pseudotyped Vesicular Stomatitis Virus-Severe Acute Respiratory Syndrome-Coronavirus-2 Spike for the Study of Variants, Vaccines, and Therapeutics Against Coronavirus Disease 2019.Front Microbiol. 2022 Jan 14;12:817200. doi: 10.3389/fmicb.2021.817200. eCollection 2021. Front Microbiol. 2022. PMID: 35095820 Free PMC article. Review.
-
Rapid Detection of Viral Envelope Lipids Using Lithium Adducts and AP-MALDI High-Resolution Mass Spectrometry.J Am Soc Mass Spectrom. 2021 Sep 1;32(9):2322-2333. doi: 10.1021/jasms.1c00058. Epub 2021 Apr 22. J Am Soc Mass Spectrom. 2021. PMID: 33886294 Free PMC article.
-
Unraveling dual fusion mechanisms in BmNPV GP64: critical roles of CARC motifs and signal peptide retention.J Virol. 2025 Jan 31;99(1):e0151124. doi: 10.1128/jvi.01511-24. Epub 2024 Nov 27. J Virol. 2025. PMID: 39601591 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
