

Functional mutants of Azospirillum brasilense elicit beneficial physiological and metabolic responses in Zea mays contributing to increased host iron assimilation
- PMID: 33408368
- PMCID: PMC8115672
- DOI: 10.1038/s41396-020-00866-x
Functional mutants of Azospirillum brasilense elicit beneficial physiological and metabolic responses in Zea mays contributing to increased host iron assimilation
Abstract
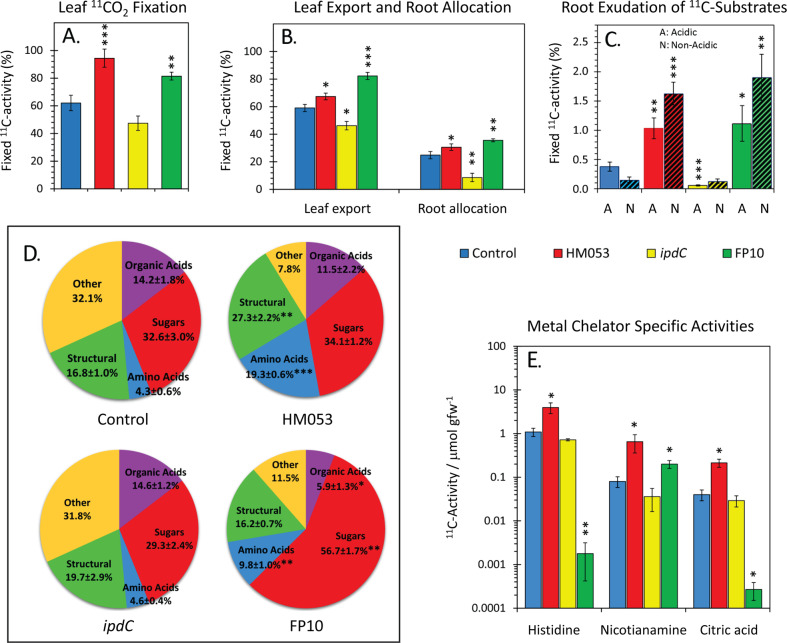
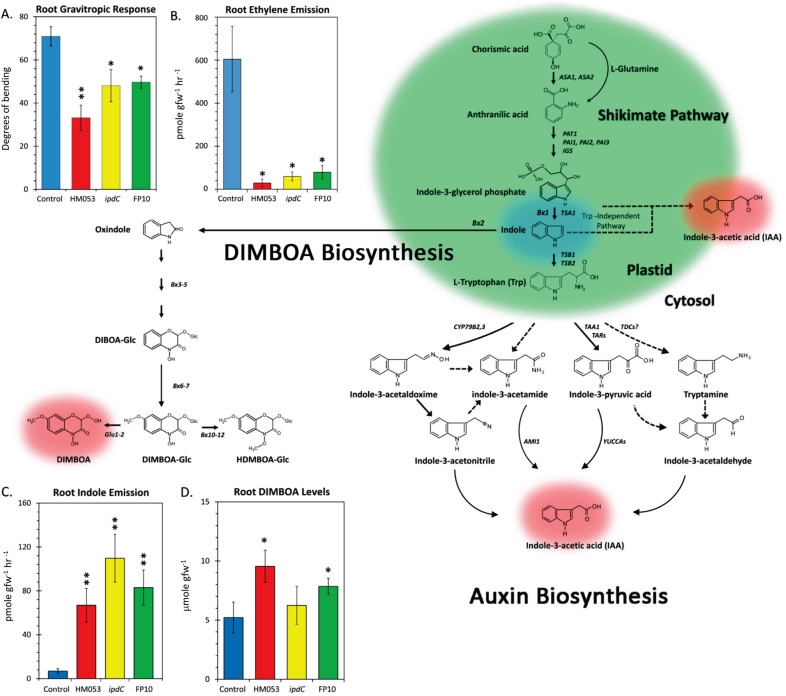
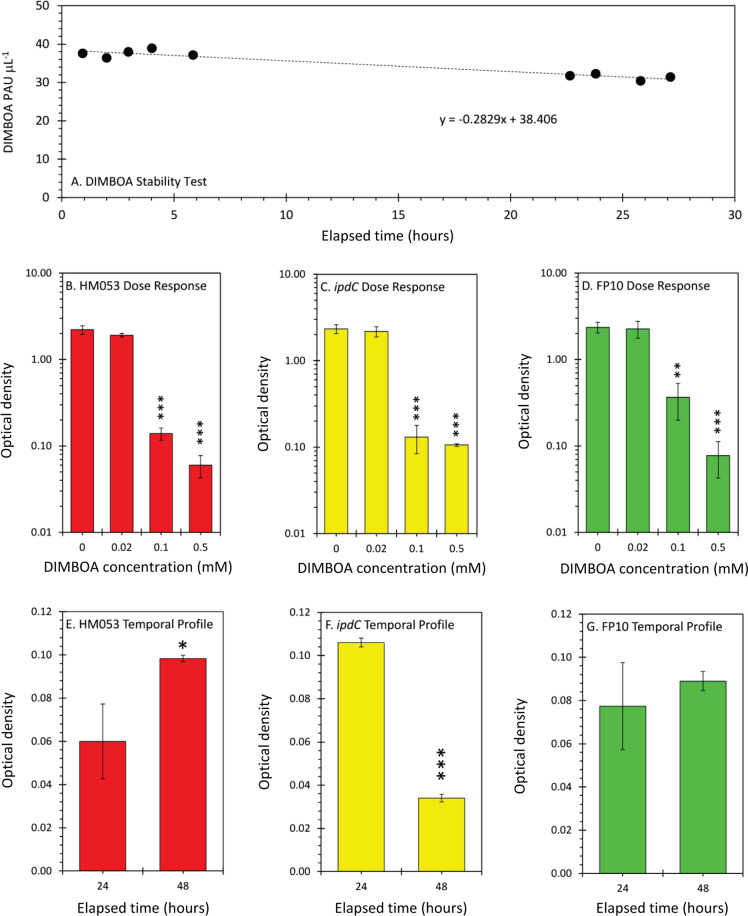
Iron (Fe), an essential element for plant growth, is abundant in soil but with low bioavailability. Thus, plants developed specialized mechanisms to sequester the element. Beneficial microbes have recently become a favored method to promote plant growth through increased uptake of essential micronutrients, like Fe, yet little is known of their mechanisms of action. Functional mutants of the epiphytic bacterium Azospirillum brasilense, a prolific grass-root colonizer, were used to examine mechanisms for promoting iron uptake in Zea mays. Mutants included HM053, FP10, and ipdC, which have varying capacities for biological nitrogen fixation and production of the plant hormone auxin. Using radioactive iron-59 tracing and inductively coupled plasma mass spectrometry, we documented significant differences in host uptake of Fe2+/3+ correlating with mutant biological function. Radioactive carbon-11, administered to plants as 11CO2, provided insights into shifts in host usage of 'new' carbon resources in the presence of these beneficial microbes. Of the mutants examined, HM053 exhibited the greatest influence on host Fe uptake with increased plant allocation of 11C-resources to roots where they were transformed and exuded as 11C-acidic substrates to aid in Fe-chelation, and increased C-11 partitioning into citric acid, nicotianamine and histidine to aid in the in situ translocation of Fe once assimilated.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures



Similar articles
-
Azospirillum brasilense Bacteria Promotes Mn2+ Uptake in Maize with Benefits to Leaf Photosynthesis.Microorganisms. 2022 Jun 25;10(7):1290. doi: 10.3390/microorganisms10071290. Microorganisms. 2022. PMID: 35889009 Free PMC article.
-
Studies Using Mutant Strains of Azospirillum brasilense Reveal That Atmospheric Nitrogen Fixation and Auxin Production Are Light Dependent Processes.Microorganisms. 2023 Jun 30;11(7):1727. doi: 10.3390/microorganisms11071727. Microorganisms. 2023. PMID: 37512900 Free PMC article.
-
Plant-Growth-Promoting Bacteria Can Impact Zinc Uptake in Zea mays: An Examination of the Mechanisms of Action Using Functional Mutants of Azospirillum brasilense.Microorganisms. 2021 May 6;9(5):1002. doi: 10.3390/microorganisms9051002. Microorganisms. 2021. PMID: 34066521 Free PMC article.
-
Key physiological properties contributing to rhizosphere adaptation and plant growth promotion abilities of Azospirillum brasilense.FEMS Microbiol Lett. 2012 Jan;326(2):99-108. doi: 10.1111/j.1574-6968.2011.02407.x. Epub 2011 Oct 3. FEMS Microbiol Lett. 2012. PMID: 22092983 Review.
-
Azospirillum, a free-living nitrogen-fixing bacterium closely associated with grasses: genetic, biochemical and ecological aspects.FEMS Microbiol Rev. 2000 Oct;24(4):487-506. doi: 10.1111/j.1574-6976.2000.tb00552.x. FEMS Microbiol Rev. 2000. PMID: 10978548 Review.
Cited by
-
Importance of the Rhizosphere Microbiota in Iron Biofortification of Plants.Front Plant Sci. 2021 Dec 3;12:744445. doi: 10.3389/fpls.2021.744445. eCollection 2021. Front Plant Sci. 2021. PMID: 34925398 Free PMC article. Review.
-
Azospirillum brasilense Bacteria Promotes Mn2+ Uptake in Maize with Benefits to Leaf Photosynthesis.Microorganisms. 2022 Jun 25;10(7):1290. doi: 10.3390/microorganisms10071290. Microorganisms. 2022. PMID: 35889009 Free PMC article.
-
Studies Using Mutant Strains of Azospirillum brasilense Reveal That Atmospheric Nitrogen Fixation and Auxin Production Are Light Dependent Processes.Microorganisms. 2023 Jun 30;11(7):1727. doi: 10.3390/microorganisms11071727. Microorganisms. 2023. PMID: 37512900 Free PMC article.
-
Plant-Growth-Promoting Bacteria Can Impact Zinc Uptake in Zea mays: An Examination of the Mechanisms of Action Using Functional Mutants of Azospirillum brasilense.Microorganisms. 2021 May 6;9(5):1002. doi: 10.3390/microorganisms9051002. Microorganisms. 2021. PMID: 34066521 Free PMC article.
-
Identification of Quantitative Trait Loci Associated With Iron Deficiency Tolerance in Maize.Front Plant Sci. 2022 Apr 14;13:805247. doi: 10.3389/fpls.2022.805247. eCollection 2022. Front Plant Sci. 2022. PMID: 35498718 Free PMC article.
References
-
- Verhagen BW, Glazebrook J, Zhu T, Chang HS, van Loon LC, Pieterse CM. The transcriptome of rhizobacteria-induced systemic resistance in Arabidopsis. Mol Plant Microbe Inter. 2004;17:895–908. - PubMed
-
- Richardson A, Barea J-M, McNeill A, Prigent-Combaret C. Acquisition of phosphorus and nitrogen in the rhizosphere and plant growth promotion by microorganisms. Plant Soil. 2009;321:305–39.
-
- Fibach-Paldi S, Burdman S, Okon Y. Key physiological properties contributing to rhizosphere adaptation and plant growth promotion abilities of Azospirillum brasilense. FEMS Microbiol Lett. 2012;326:99e108. - PubMed
-
- Pankievicz VCS, Amaral FP, Santos KFD, Agtuca B, Xu Y, Schueller MJ, et al. Robust biological nitrogen fixation in a model grass-bacterial association. Plant J. 2015;81:907–19. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
