

A STAT3 inhibitor ameliorates CNS autoimmunity by restoring Teff:Treg balance
- PMID: 33411696
- PMCID: PMC7934926
- DOI: 10.1172/jci.insight.142376
A STAT3 inhibitor ameliorates CNS autoimmunity by restoring Teff:Treg balance
Abstract
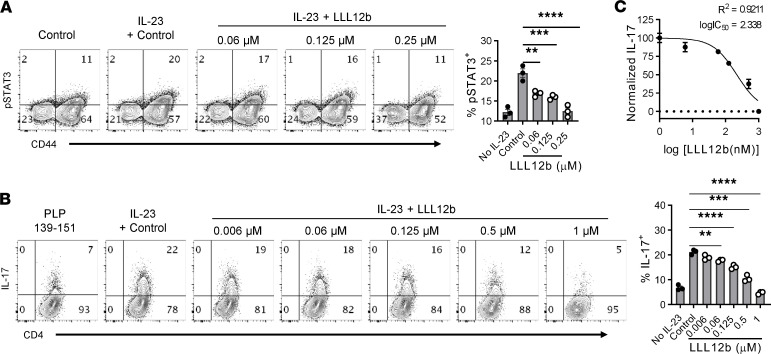
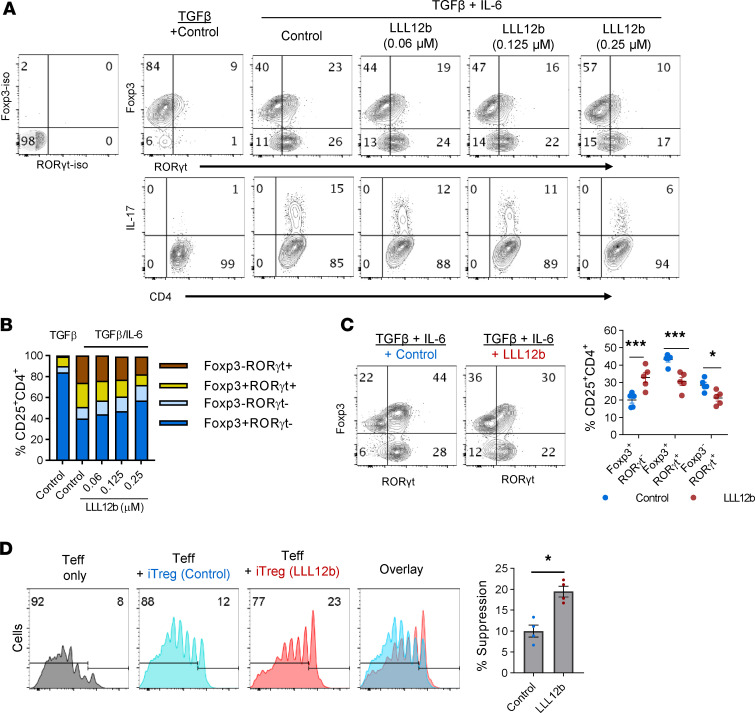
Reestablishing an appropriate balance between T effector cells (Teff) and Tregs is essential for correcting autoimmunity. Multiple sclerosis (MS) is an immune-mediated chronic CNS disease characterized by neuroinflammation, demyelination, and neuronal degeneration, in which the Teff:Treg balance is skewed toward pathogenic Teffs Th1 and Th17 cells. STAT3 is a key regulator of Teff:Treg balance. Using the structure-based design, we have developed a potentially novel small-molecule prodrug LLL12b that specifically inhibits STAT3 and suppresses Th17 differentiation and expansion. Moreover, LLL12b regulates the fate decision between Th17 and Tregs in an inflammatory environment, shifting Th17:Treg balance toward Tregs and favoring the resolution of inflammation. Therapeutic administration of LLL12b after disease onset significantly suppresses disease progression in adoptively transferred, chronic, and relapsing-remitting experimental autoimmune encephalomyelitis. Disease relapses were also significantly suppressed by LLL12b given during the remission phase. Additionally, LLL12b shifts Th17:Treg balance of CD4+ T cells from MS patients toward Tregs and increases Teff sensitivity to Treg-mediated suppression. These data suggest that selective inhibition of STAT3 by the small molecule LLL12b recalibrates the effector and regulatory arms of CD4+ T responses, representing a potentially clinically translatable therapeutic strategy for MS.
Keywords: Autoimmune diseases; Autoimmunity; Demyelinating disorders; Drug therapy; Therapeutics.
Conflict of interest statement
Figures



References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
