

An influenza HA stalk reactive polymeric IgA antibody exhibits anti-viral function regulated by binary interaction between HA and the antibody
- PMID: 33412571
- PMCID: PMC7790537
- DOI: 10.1371/journal.pone.0245244
An influenza HA stalk reactive polymeric IgA antibody exhibits anti-viral function regulated by binary interaction between HA and the antibody
Abstract
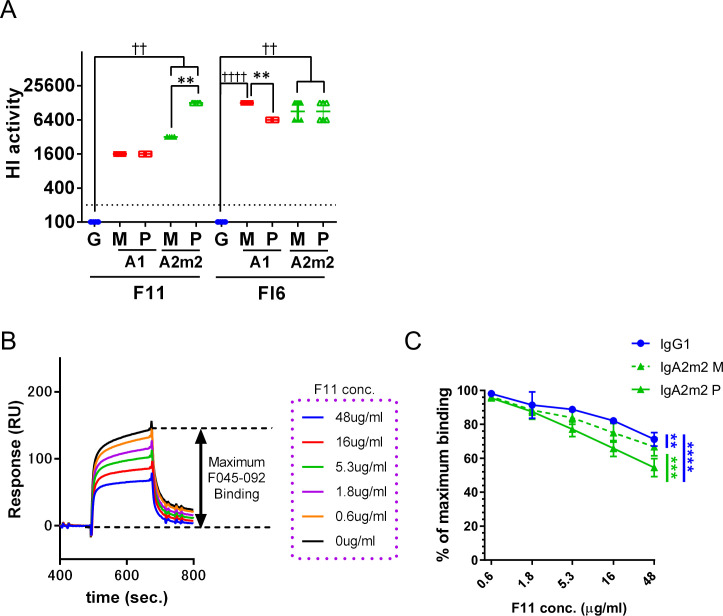
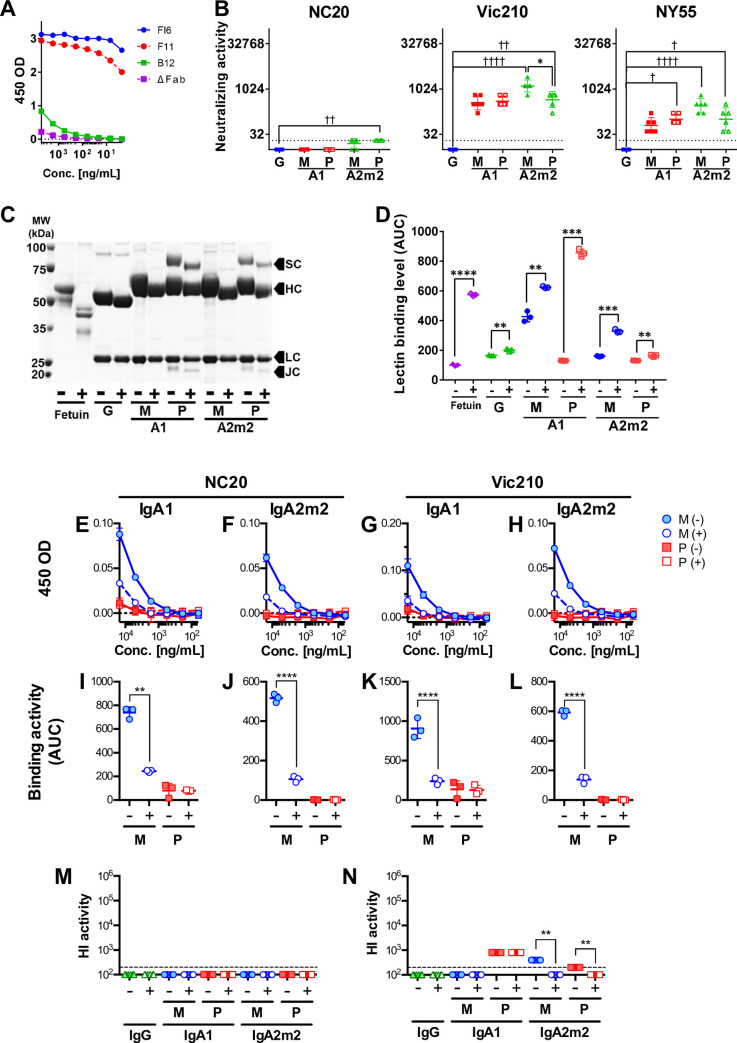
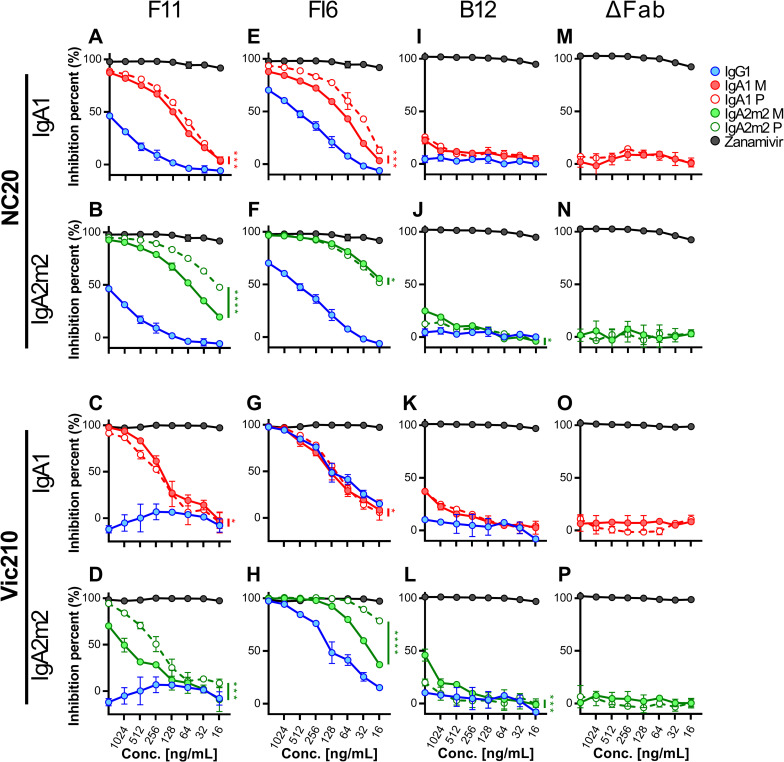
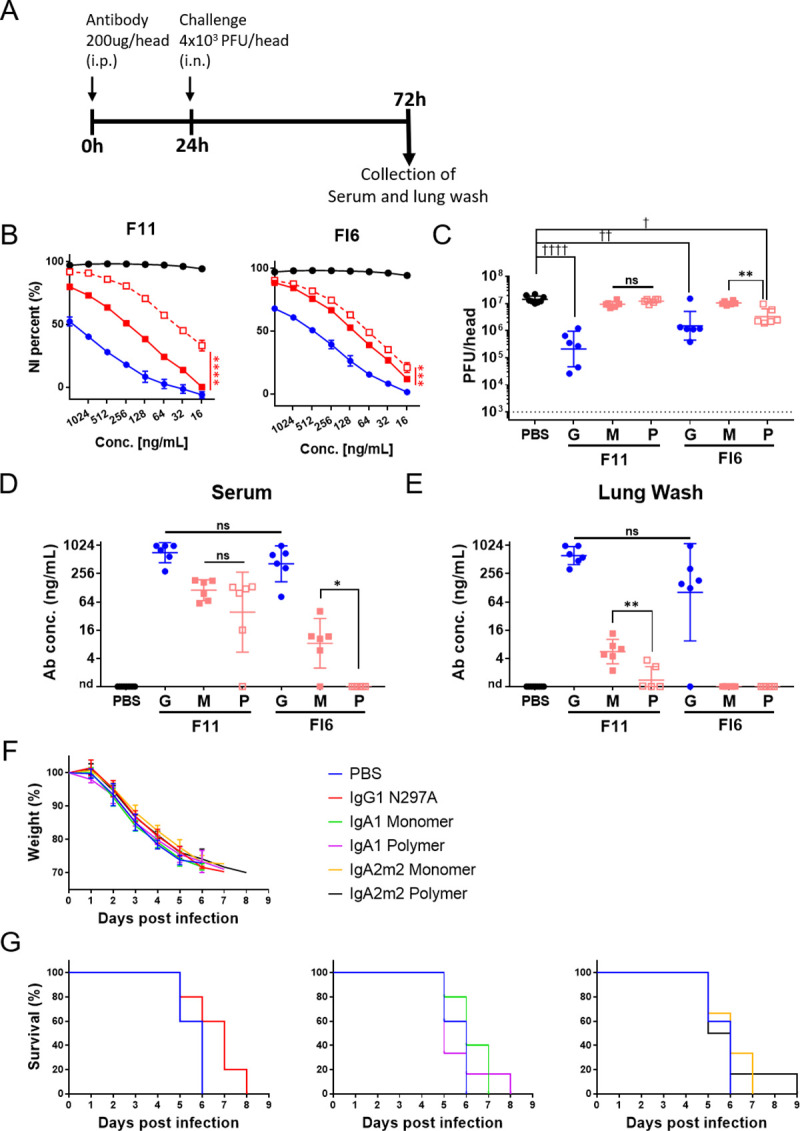
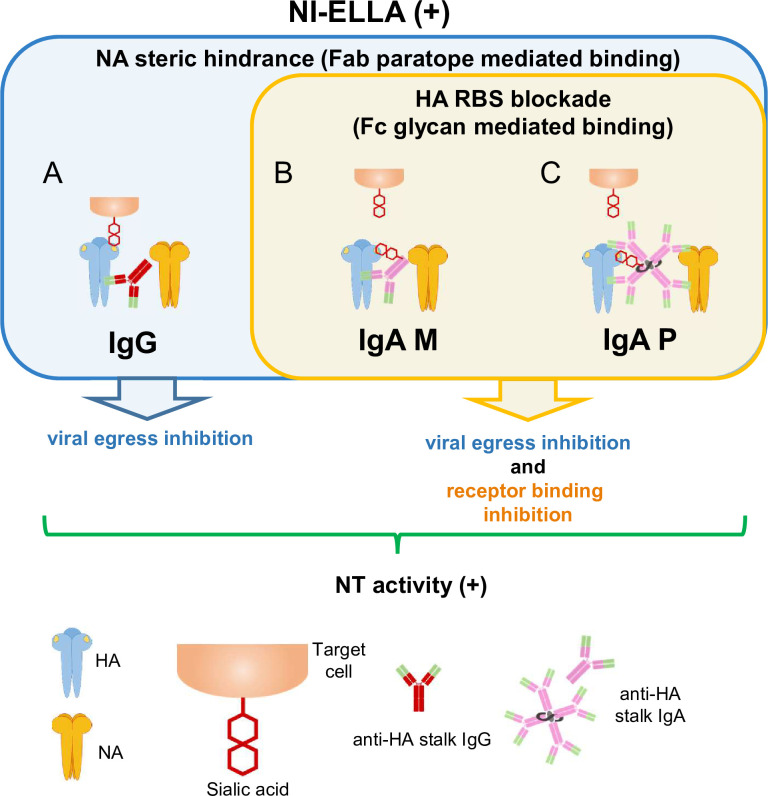
IgA antibodies, which are secreted onto the mucosal surface as secretory IgA antibodies (SIgAs), play an important role in preventing influenza virus infection. A recent study reported that anti-hemagglutinin (HA) head-targeting antibodies increase anti-viral functions such as hemagglutination inhibition (HI) and virus neutralization (NT), in addition to HA binding activity (reactivity) via IgA polymerization. However, the functional properties of anti-viral IgA antibodies with mechanisms of action distinct from those of anti-HA head-targeting antibodies remain elusive. Here, we characterized the functional properties of IgG, monomeric IgA, and polymeric IgA anti-HA stalk-binding clones F11 and FI6, and B12 (a low affinity anti-HA stalk clone), as well as Fab-deficient (ΔFab) IgA antibodies. We found that IgA polymerization impacts the functional properties of anti-HA stalk antibodies. Unlike anti-HA head antibodies, the anti-viral functions of anti-HA stalk antibodies were not simply enhanced by IgA polymerization. The data suggest that two modes of binding (Fab paratope-mediated binding to the HA stalk, and IgA Fc glycan-mediated binding to the HA receptor binding site (RBS)) occur during interaction between anti-stalk HA IgA antibodies and HA. In situations where Fab paratope-mediated binding to the HA stalk exceeded IgA Fc glycan-mediated binding to HA RBS, IgA polymerization increased anti-viral functions. By contrast, when IgA Fc glycan-mediated binding to the HA RBS was dominant, anti-viral activity will fall upon IgA polymerization. In summary, the results suggest that coordination between these two independent binding modules determines whether IgA polymerization has a negative or positive effect on the anti-viral functions of anti-HA stalk IgA antibodies.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
Similar articles
-
Structure-Guided Creation of an Anti-HA Stalk Antibody F11 Derivative That Neutralizes Both F11-Sensitive and -Resistant Influenza A(H1N1)pdm09 Viruses.Viruses. 2021 Aug 31;13(9):1733. doi: 10.3390/v13091733. Viruses. 2021. PMID: 34578314 Free PMC article.
-
Potential Role of Nonneutralizing IgA Antibodies in Cross-Protective Immunity against Influenza A Viruses of Multiple Hemagglutinin Subtypes.J Virol. 2020 Jun 1;94(12):e00408-20. doi: 10.1128/JVI.00408-20. Print 2020 Jun 1. J Virol. 2020. PMID: 32269119 Free PMC article.
-
Extending the Stalk Enhances Immunogenicity of the Influenza Virus Neuraminidase.J Virol. 2019 Aug 28;93(18):e00840-19. doi: 10.1128/JVI.00840-19. Print 2019 Sep 15. J Virol. 2019. PMID: 31375573 Free PMC article.
-
Broadly neutralizing anti-influenza virus antibodies: enhancement of neutralizing potency in polyclonal mixtures and IgA backbones.J Virol. 2015 Apr;89(7):3610-8. doi: 10.1128/JVI.03099-14. Epub 2015 Jan 14. J Virol. 2015. PMID: 25589655 Free PMC article.
-
IgA tetramerization improves target breadth but not peak potency of functionality of anti-influenza virus broadly neutralizing antibody.PLoS Pathog. 2019 Jan 3;15(1):e1007427. doi: 10.1371/journal.ppat.1007427. eCollection 2019 Jan. PLoS Pathog. 2019. PMID: 30605488 Free PMC article.
Cited by
-
Induction of Tier 2 HIV-Neutralizing IgA Antibodies in Rhesus Macaques Vaccinated with BG505.664 SOSIP.Vaccines (Basel). 2024 Dec 10;12(12):1386. doi: 10.3390/vaccines12121386. Vaccines (Basel). 2024. PMID: 39772048 Free PMC article.
-
Cis-Allosteric Regulation of HIV-1 Reverse Transcriptase by Integrase.Viruses. 2022 Dec 21;15(1):31. doi: 10.3390/v15010031. Viruses. 2022. PMID: 36680070 Free PMC article.
-
Structure-Guided Creation of an Anti-HA Stalk Antibody F11 Derivative That Neutralizes Both F11-Sensitive and -Resistant Influenza A(H1N1)pdm09 Viruses.Viruses. 2021 Aug 31;13(9):1733. doi: 10.3390/v13091733. Viruses. 2021. PMID: 34578314 Free PMC article.
References
-
- Tamura S, Funato H, Hirabayashi Y, Kikuta K, Suzuki Y, Nagamine T, et al. Functional role of respiratory tract haemagglutinin-specific IgA antibodies in protection against influenza. Vaccine. 1990;8: 479–85. Available: http://www.ncbi.nlm.nih.gov/pubmed/2251874 10.1016/0264-410x(90)90250-p - DOI - PubMed
-
- Waldman RH, Wood SH, Torres EJ, Small PA. Influenza antibody response following aerosal administration of inactivated virus. Am J Epidemiol. 1970;91: 574–85. Available: http://www.ncbi.nlm.nih.gov/pubmed/5445644 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
