

Tubulin glycylation controls axonemal dynein activity, flagellar beat, and male fertility
- PMID: 33414192
- PMCID: PMC7612590
- DOI: 10.1126/science.abd4914
Tubulin glycylation controls axonemal dynein activity, flagellar beat, and male fertility
Abstract
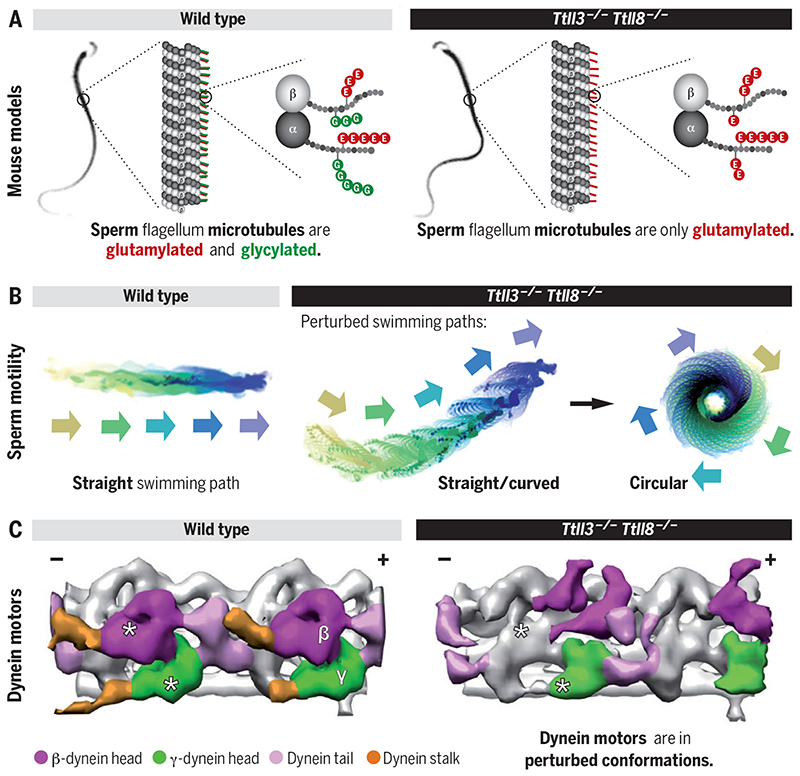
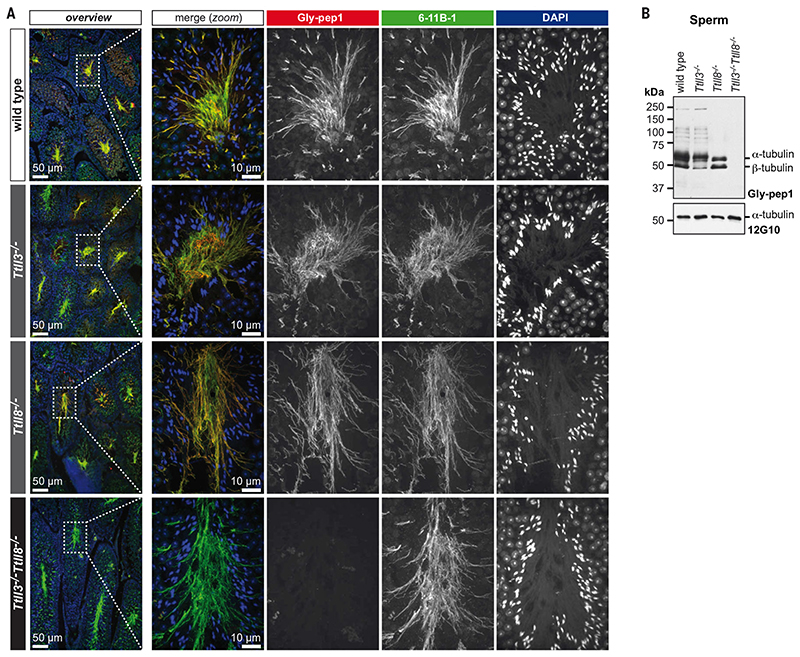
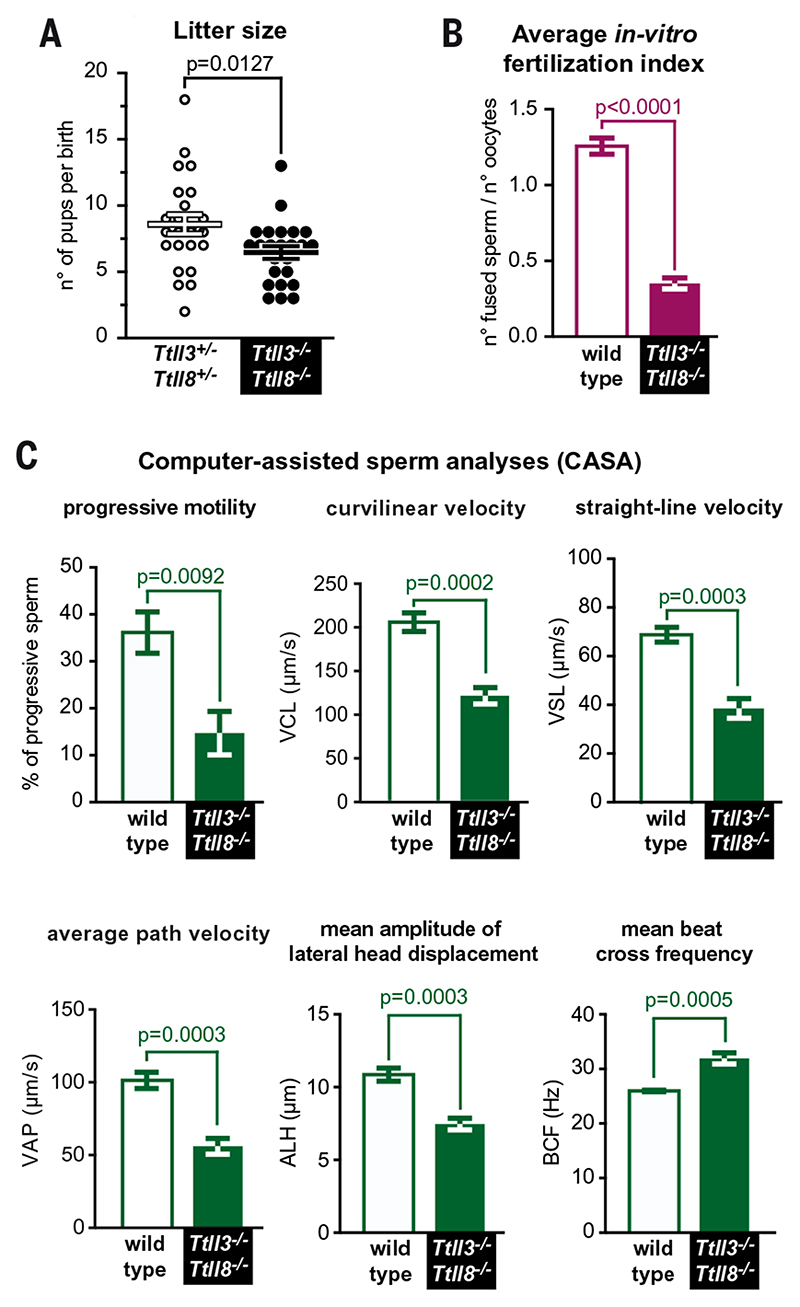
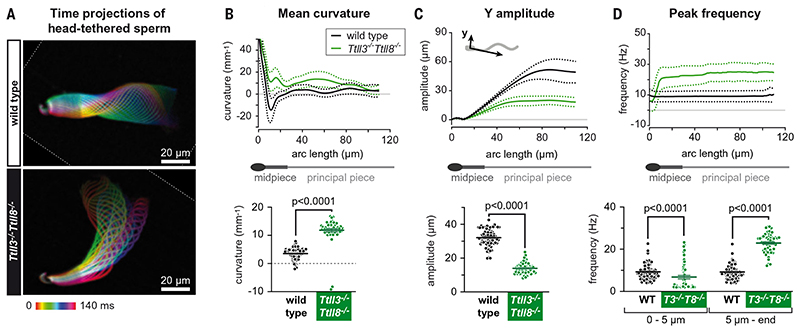
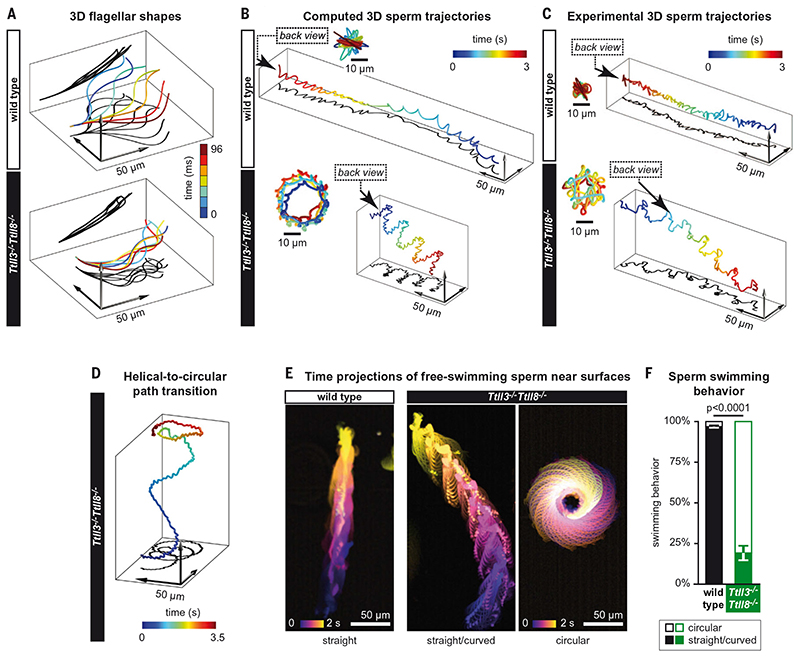
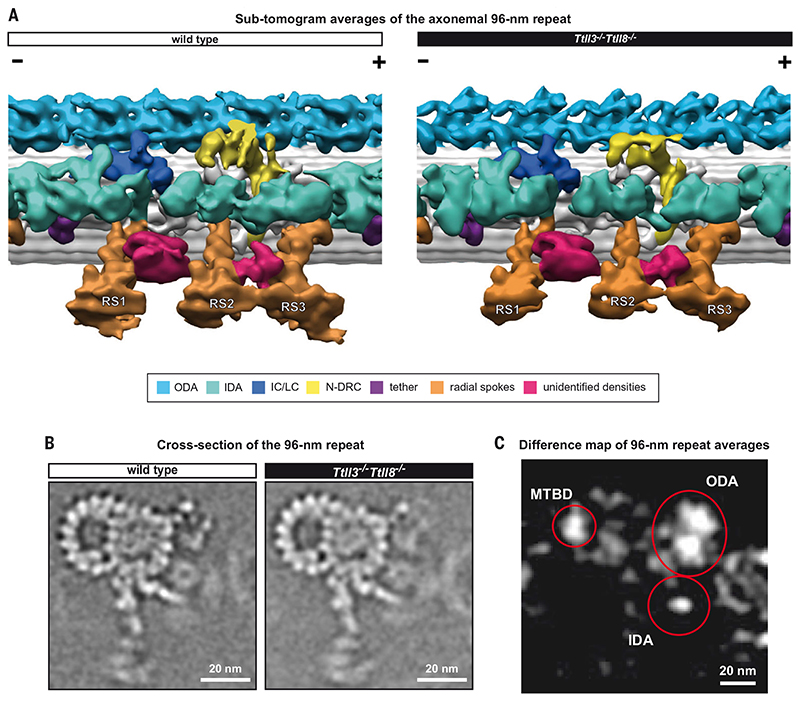
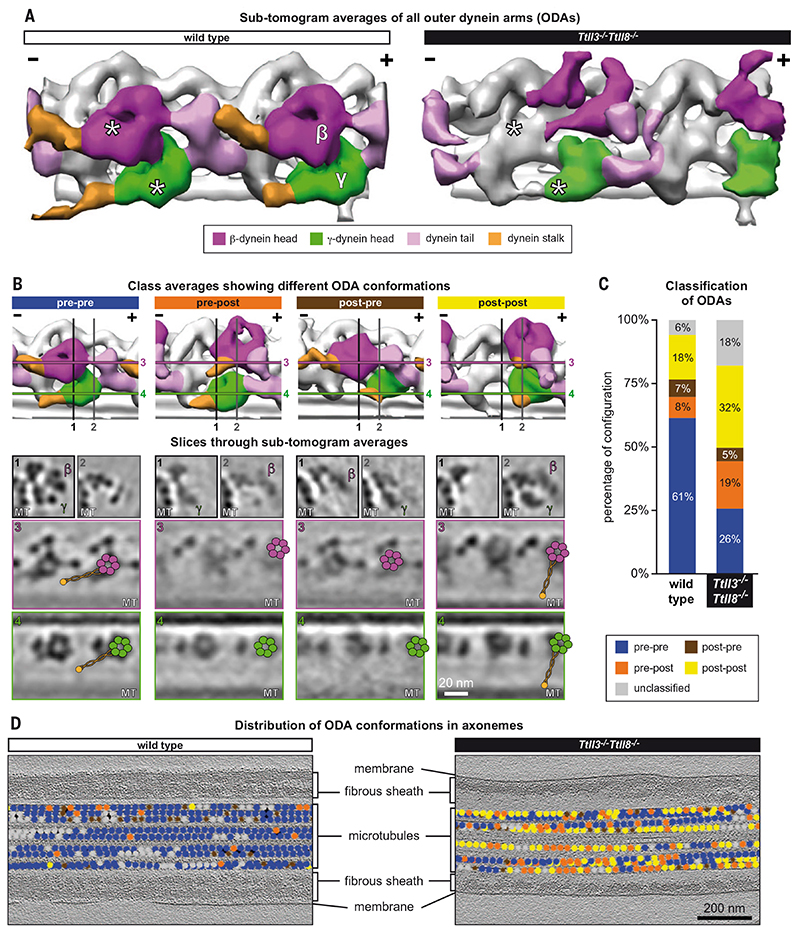
Posttranslational modifications of the microtubule cytoskeleton have emerged as key regulators of cellular functions, and their perturbations have been linked to a growing number of human pathologies. Tubulin glycylation modifies microtubules specifically in cilia and flagella, but its functional and mechanistic roles remain unclear. In this study, we generated a mouse model entirely lacking tubulin glycylation. Male mice were subfertile owing to aberrant beat patterns of their sperm flagella, which impeded the straight swimming of sperm cells. Using cryo-electron tomography, we showed that lack of glycylation caused abnormal conformations of the dynein arms within sperm axonemes, providing the structural basis for the observed dysfunction. Our findings reveal the importance of microtubule glycylation for controlled flagellar beating, directional sperm swimming, and male fertility.
Copyright © 2021, American Association for the Advancement of Science.
Conflict of interest statement
Figures
Comment in
-
Sperm going in circles.Nat Rev Mol Cell Biol. 2021 Apr;22(4):242-243. doi: 10.1038/s41580-021-00345-x. Nat Rev Mol Cell Biol. 2021. PMID: 33558683 No abstract available.
Similar articles
-
Tubulin glycylation controls ciliary motility through modulation of outer-arm dyneins.Mol Biol Cell. 2024 Jul 1;35(7):ar90. doi: 10.1091/mbc.E24-04-0154. Epub 2024 May 17. Mol Biol Cell. 2024. PMID: 38758663 Free PMC article.
-
Defects in the cytoplasmic assembly of axonemal dynein arms cause morphological abnormalities and dysmotility in sperm cells leading to male infertility.PLoS Genet. 2021 Feb 26;17(2):e1009306. doi: 10.1371/journal.pgen.1009306. eCollection 2021 Feb. PLoS Genet. 2021. PMID: 33635866 Free PMC article.
-
The motility of axonemal dynein is regulated by the tubulin code.Biophys J. 2014 Dec 16;107(12):2872-2880. doi: 10.1016/j.bpj.2014.10.061. Biophys J. 2014. PMID: 25658008 Free PMC article.
-
Posttranslational Modifications of Tubulin and Cilia.Cold Spring Harb Perspect Biol. 2017 Jun 1;9(6):a028159. doi: 10.1101/cshperspect.a028159. Cold Spring Harb Perspect Biol. 2017. PMID: 28003186 Free PMC article. Review.
-
Ciliary and flagellar structure and function--their regulations by posttranslational modifications of axonemal tubulin.Int Rev Cell Mol Biol. 2012;294:133-70. doi: 10.1016/B978-0-12-394305-7.00003-3. Int Rev Cell Mol Biol. 2012. PMID: 22364873 Review.
Cited by
-
Cilia at the crossroad: convergence of regulatory mechanisms to govern cilia dynamics during cell signaling and the cell cycle.Cell Biosci. 2025 Jun 7;15(1):81. doi: 10.1186/s13578-025-01403-z. Cell Biosci. 2025. PMID: 40483459 Free PMC article. Review.
-
Cylicins are a structural component of the sperm calyx being indispensable for male fertility in mice and human.Elife. 2023 Nov 28;12:RP86100. doi: 10.7554/eLife.86100. Elife. 2023. PMID: 38013430 Free PMC article.
-
Epigenetically Mediated Ciliogenesis and Cell Cycle Regulation, and Their Translational Potential.Cells. 2021 Jul 2;10(7):1662. doi: 10.3390/cells10071662. Cells. 2021. PMID: 34359832 Free PMC article. Review.
-
DOT1L regulates chromatin reorganization and gene expression during sperm differentiation.EMBO Rep. 2023 Jun 5;24(6):e56316. doi: 10.15252/embr.202256316. Epub 2023 Apr 26. EMBO Rep. 2023. PMID: 37099396 Free PMC article.
-
Ciliary length regulation by intraflagellar transport in zebrafish.Elife. 2024 Dec 13;13:RP93168. doi: 10.7554/eLife.93168. Elife. 2024. PMID: 39671305 Free PMC article.
References
-
- Bré MH, et al. Axonemal tubulin polyglycylation probed with two monoclonal antibodies: Widespread evolutionary distribution, appearance during spermatozoan maturation and possible function in motility. J Cell Sci. 1996;109:727–738. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
