

The Highly Conservative Cysteine of Oncomodulin as a Feasible Redox Sensor
- PMID: 33419032
- PMCID: PMC7825312
- DOI: 10.3390/biom11010066
The Highly Conservative Cysteine of Oncomodulin as a Feasible Redox Sensor
Abstract
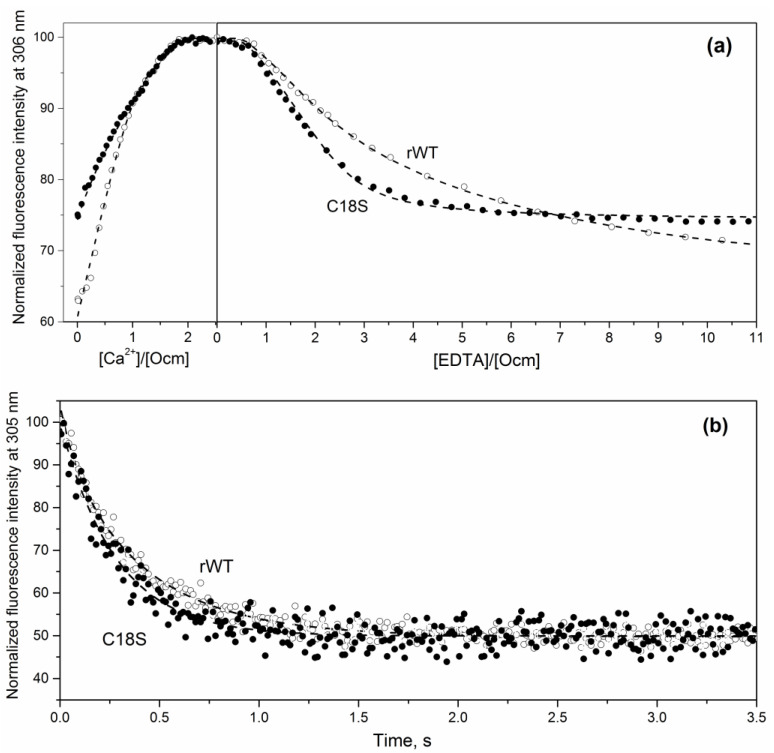
Oncomodulin (Ocm), or parvalbumin β, is an 11-12 kDa Ca2+-binding protein found inside and outside of vertebrate cells, which regulates numerous processes via poorly understood mechanisms. Ocm consists of two active Ca2+-specific domains of the EF-hand type ("helix-loop-helix" motif), covered by an EF-hand domain with inactive EF-hand loop, which contains a highly conservative cysteine with unknown function. In this study, we have explored peculiarities of the microenvironment of the conservative Cys18 of recombinant rat Ocm (rWT Ocm), redox properties of this residue, and structural/functional sensitivity of rWT Ocm to the homologous C18S substitution. We have found that pKa of the Cys18 thiol lays beyond the physiological pH range. The measurement of redox dependence of rWT Ocm thiol-disulfide equilibrium (glutathione redox pair) showed that redox potential of Cys18 for the metal-free and Ca2+-loaded protein is of -168 mV and -176 mV, respectively. Therefore, the conservative thiol of rWT Ocm is prone to disulfide dimerization under physiological redox conditions. The C18S substitution drastically reduces α-helices content of the metal-free and Mg2+-bound Ocm, increases solvent accessibility of its hydrophobic residues, eliminates the cooperative thermal transition in the apo-protein, suppresses Ca2+/Mg2+ affinity of the EF site, and accelerates Ca2+ dissociation from Ocm. The distinct structural and functional consequences of the minor structural modification of Cys18 indicate its possible redox sensory function. Since some other EF-hand proteins also contain a conservative redox-sensitive cysteine located in an inactive EF-hand loop, it is reasonable to suggest that in the course of evolution, some of the EF-hands attained redox sensitivity at the expense of the loss of their Ca2+ affinity.
Keywords: EF-hand; calcium-binding protein; cysteine; disulfide dimerization; oncomodulin; parvalbumin; protein stability; redox potential; redox sensor; thiol oxidation.
Conflict of interest statement
The authors declare no conflict of interest.
Figures




Similar articles
-
Strontium Binding to α-Parvalbumin, a Canonical Calcium-Binding Protein of the "EF-Hand" Family.Biomolecules. 2021 Aug 5;11(8):1158. doi: 10.3390/biom11081158. Biomolecules. 2021. PMID: 34439824 Free PMC article.
-
Remodeling of the AB site of rat parvalbumin and oncomodulin into a canonical EF-hand.Eur J Biochem. 1999 Sep;264(3):790-9. doi: 10.1046/j.1432-1327.1999.00650.x. Eur J Biochem. 1999. PMID: 10491125
-
Introduction of a fifth carboxylate ligand heightens the affinity of the oncomodulin CD and EF sites for Ca2+.Biochemistry. 1996 May 7;35(18):5856-69. doi: 10.1021/bi952184d. Biochemistry. 1996. PMID: 8639547
-
Structures and metal-ion-binding properties of the Ca2+-binding helix-loop-helix EF-hand motifs.Biochem J. 2007 Jul 15;405(2):199-221. doi: 10.1042/BJ20070255. Biochem J. 2007. PMID: 17590154 Review.
-
Functional diversity of cysteine residues in proteins and unique features of catalytic redox-active cysteines in thiol oxidoreductases.Mol Cells. 2008 Sep 30;26(3):228-35. Epub 2008 Jul 23. Mol Cells. 2008. PMID: 18648218 Free PMC article. Review.
Cited by
-
Parvalbumin Role in Epilepsy and Psychiatric Comorbidities: From Mechanism to Intervention.Front Integr Neurosci. 2022 Feb 17;16:765324. doi: 10.3389/fnint.2022.765324. eCollection 2022. Front Integr Neurosci. 2022. PMID: 35250498 Free PMC article. Review.
-
Comprehensive Sequence Analysis of Parvalbumins in Fish and Their Comparison with Parvalbumins in Tetrapod Species.Biology (Basel). 2022 Nov 25;11(12):1713. doi: 10.3390/biology11121713. Biology (Basel). 2022. PMID: 36552222 Free PMC article.
-
Disulfide Dimerization of Neuronal Calcium Sensor-1: Implications for Zinc and Redox Signaling.Int J Mol Sci. 2021 Nov 22;22(22):12602. doi: 10.3390/ijms222212602. Int J Mol Sci. 2021. PMID: 34830487 Free PMC article.
References
-
- MacManus J.P. Occurrence of a low-molecular-weight calcium-binding protein in neoplastic liver. Cancer Res. 1979;39:3000–3005. - PubMed
-
- Permyakov E.A. Parvalbumin. In: Uversky V.N., editor. series Molecular Anatomy and Physiology of Proteins. Nova Science Publishers, Inc.; New York, NY, USA: 2006.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
