

Species-Specific and Distance-Dependent Dispersive Behaviour of Forisomes in Different Legume Species
- PMID: 33419062
- PMCID: PMC7825422
- DOI: 10.3390/ijms22020492
Species-Specific and Distance-Dependent Dispersive Behaviour of Forisomes in Different Legume Species
Abstract
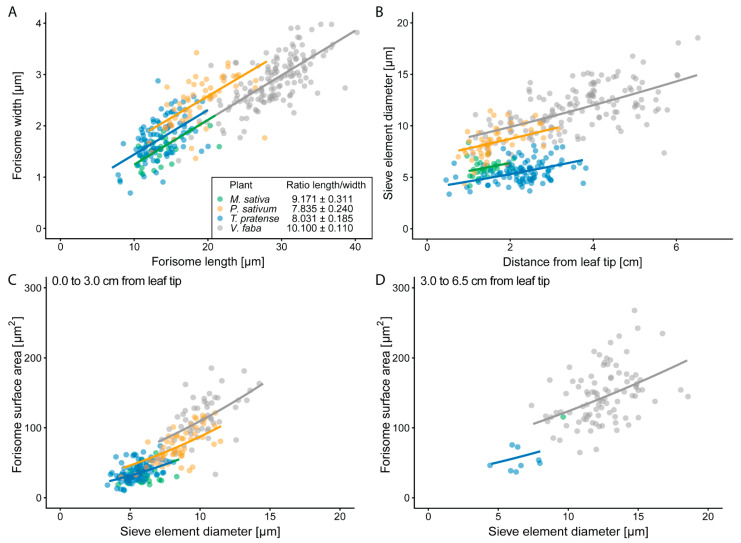
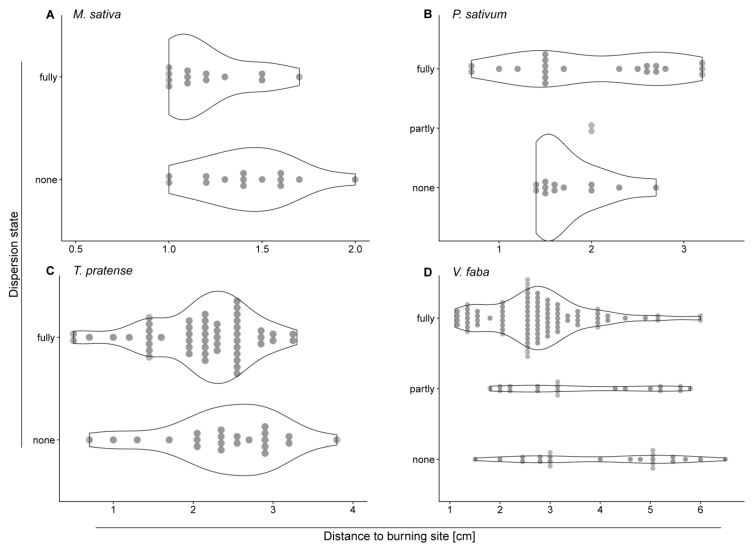
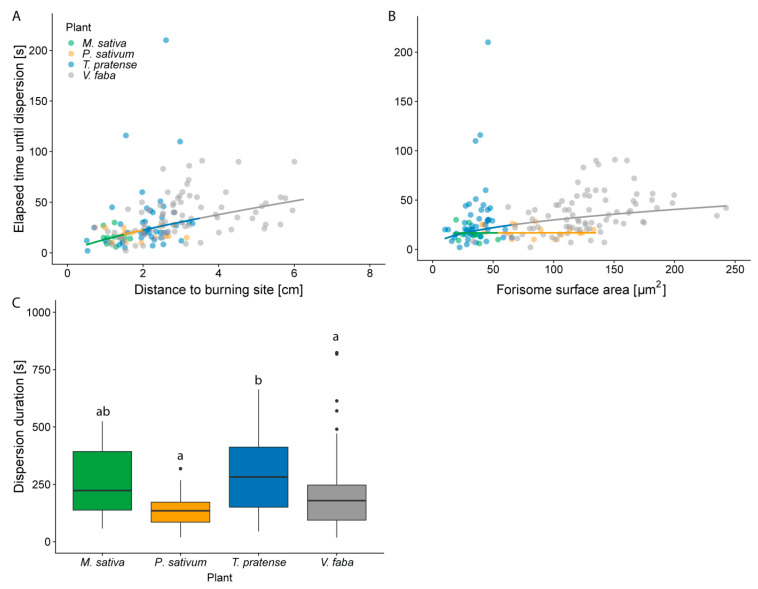
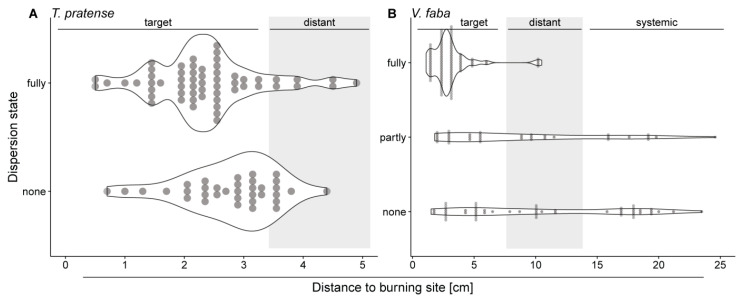
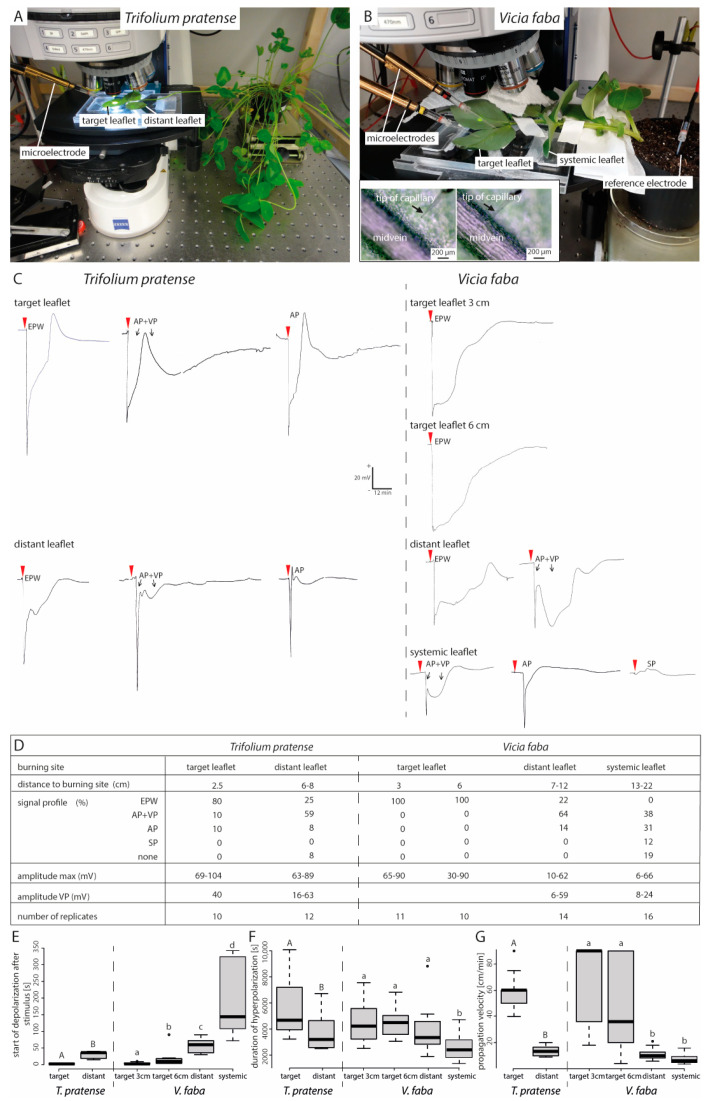
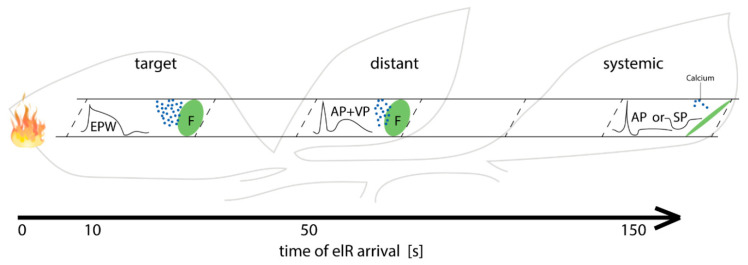
Forisomes are giant fusiform protein complexes composed of sieve element occlusion (SEO) protein monomers, exclusively found in sieve elements (SEs) of legumes. Forisomes block the phloem mass flow by a Ca2+-induced conformational change (swelling and rounding). We studied the forisome reactivity in four different legume species-Medicago sativa, Pisum sativum, Trifolium pratense and Vicia faba. Depending on the species, we found direct relationships between SE diameter, forisome surface area and distance from the leaf tip, all indicative of a developmentally tuned regulation of SE diameter and forisome size. Heat-induced forisome dispersion occurred later with increasing distance from the stimulus site. T. pratense and V. faba dispersion occurred faster for forisomes with a smaller surface area. Near the stimulus site, electro potential waves (EPWs)-overlapping action (APs), and variation potentials (VPs)-were linked with high full-dispersion rates of forisomes. Distance-associated reduction of forisome reactivity was assigned to the disintegration of EPWs into APs, VPs and system potentials (SPs). Overall, APs and SPs alone were unable to induce forisome dispersion and only VPs above a critical threshold were capable of inducing forisome reactions.
Keywords: Medicago sativa; Pisum sativum; Trifolium pratense; Vicia faba; electrophysiology; forisome; sieve-tube occlusion; systemic signalling.
Conflict of interest statement
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
Figures

Similar articles
-
Pea Aphid (Acyrthosiphon pisum) Host Races Reduce Heat-Induced Forisome Dispersion in Vicia faba and Trifolium pratense.Plants (Basel). 2023 May 6;12(9):1888. doi: 10.3390/plants12091888. Plants (Basel). 2023. PMID: 37176952 Free PMC article.
-
Comparison of intracellular location and stimulus reaction times of forisomes in sieve tubes of four legume species.Plant Signal Behav. 2018;13(8):e1503493. doi: 10.1080/15592324.2018.1503493. Epub 2018 Aug 15. Plant Signal Behav. 2018. PMID: 30111246 Free PMC article.
-
Similar Intracellular Location and Stimulus Reactivity, but Differential Mobility of Tailless (Vicia faba) and Tailed Forisomes (Phaseolus vulgaris) in Intact Sieve Tubes.PLoS One. 2015 Dec 1;10(12):e0143920. doi: 10.1371/journal.pone.0143920. eCollection 2015. PLoS One. 2015. PMID: 26624625 Free PMC article.
-
Sieve element occlusion: Interactions with phloem sap-feeding insects. A review.J Plant Physiol. 2022 Feb;269:153582. doi: 10.1016/j.jplph.2021.153582. Epub 2021 Dec 5. J Plant Physiol. 2022. PMID: 34953413 Review.
-
Calcium powered phloem protein of SEO gene family "Forisome" functions in wound sealing and act as biomimetic smart materials.Plant Signal Behav. 2014;9(9):e29438. doi: 10.4161/psb.29438. Plant Signal Behav. 2014. PMID: 25763691 Free PMC article. Review.
Cited by
-
Pea Aphid (Acyrthosiphon pisum) Host Races Reduce Heat-Induced Forisome Dispersion in Vicia faba and Trifolium pratense.Plants (Basel). 2023 May 6;12(9):1888. doi: 10.3390/plants12091888. Plants (Basel). 2023. PMID: 37176952 Free PMC article.
-
Transformation of flg22 perception into electrical signals decoded in vasculature leads to sieve tube blockage and pathogen resistance.Sci Adv. 2025 Feb 28;11(9):eads6417. doi: 10.1126/sciadv.ads6417. Epub 2025 Feb 26. Sci Adv. 2025. PMID: 40009682 Free PMC article.
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
