

N-Glycosylation as a Tool to Study Antithrombin Secretion, Conformation, and Function
- PMID: 33419227
- PMCID: PMC7825591
- DOI: 10.3390/ijms22020516
N-Glycosylation as a Tool to Study Antithrombin Secretion, Conformation, and Function
Abstract
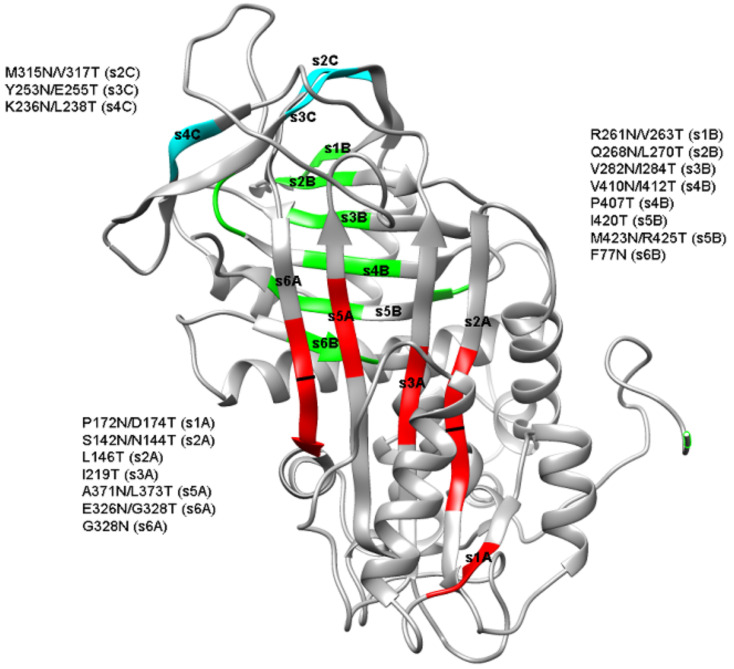
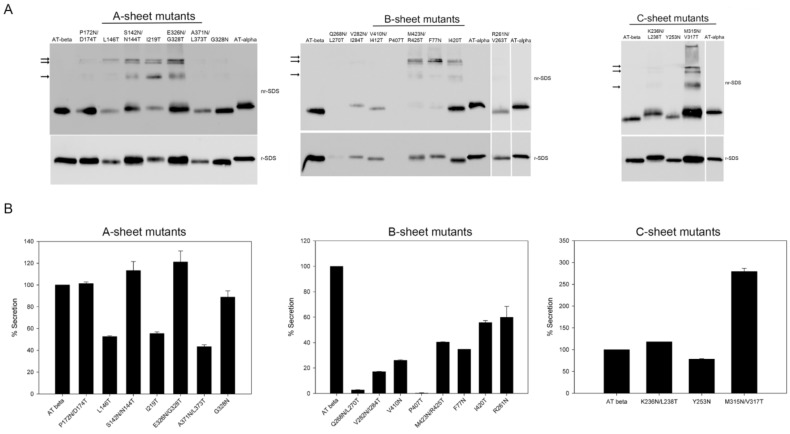
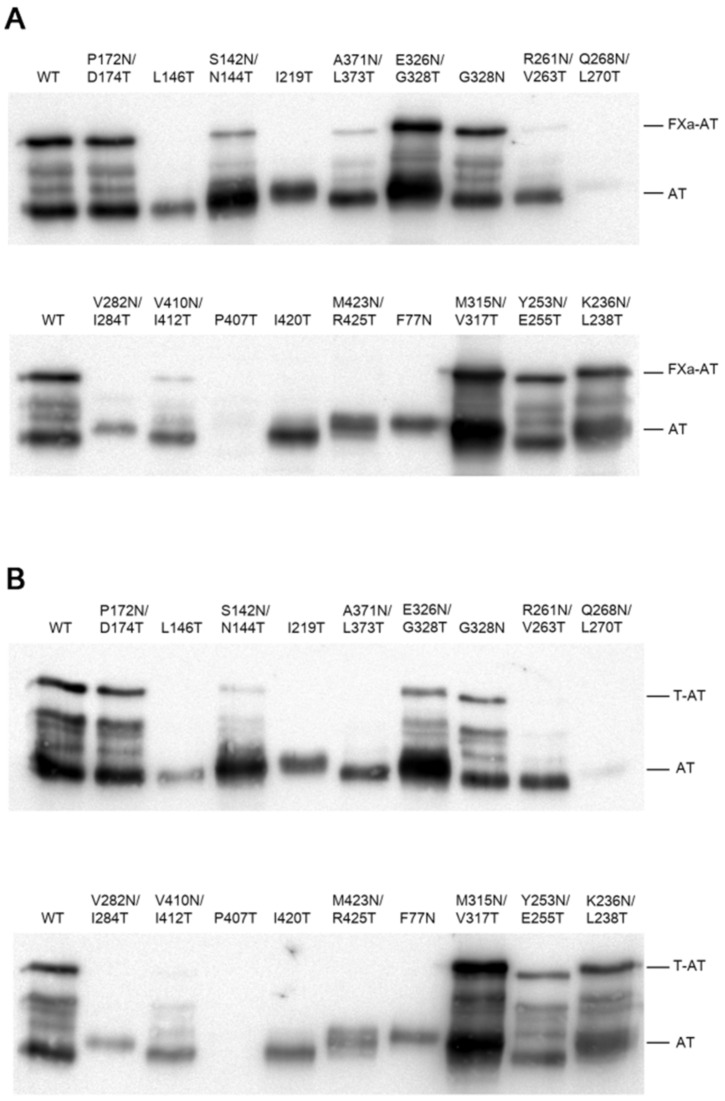
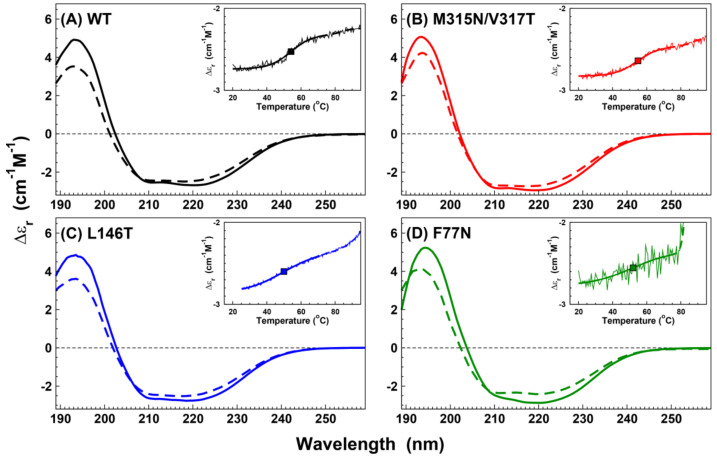
N-linked glycosylation is a crucial post-translational modification involved in protein folding, function, and clearance. N-linked glycosylation is also used therapeutically to enhance the half-lives of many proteins. Antithrombin, a serpin with four potential N-glycosylation sites, plays a pivotal role in hemostasis, wherein its deficiency significantly increases thrombotic risk. In this study, we used the introduction of N-glycosylation sites as a tool to explore what effect this glycosylation has on the protein folding, secretion, and function of this key anticoagulant. To accomplish this task, we introduced an additional N-glycosylation sequence in each strand. Interestingly, all regions that likely fold rapidly or were surrounded by lysines were not glycosylated even though an N-glycosylation sequon was present. The new sequon in the strands of the A- and B-sheets reduced secretion, and the B-sheet was more sensitive to these changes. However, the mutations in the strands of the C-sheet allowed correct folding and secretion, which resulted in functional variants. Therefore, our study revealed crucial regions for antithrombin secretion and could potentially apply to all serpins. These results could also help us understand the functional effects of natural variants causing type-I deficiencies.
Keywords: Glycosylation; antithrombin; bioengineering; coagulation; folding; serpin; thrombosis.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
Increased N-glycosylation efficiency by generation of an aromatic sequon on N135 of antithrombin.PLoS One. 2014 Dec 8;9(12):e114454. doi: 10.1371/journal.pone.0114454. eCollection 2014. PLoS One. 2014. PMID: 25485983 Free PMC article.
-
The infective polymerization of conformationally unstable antithrombin mutants may play a role in the clinical severity of antithrombin deficiency.Mol Med. 2012 Jul 18;18(1):762-70. doi: 10.2119/molmed.2012.00017. Mol Med. 2012. PMID: 22481271 Free PMC article.
-
Antithrombin Murcia (K241E) causing antithrombin deficiency: a possible role for altered glycosylation.Haematologica. 2010 Aug;95(8):1358-65. doi: 10.3324/haematol.2009.015487. Epub 2010 Apr 30. Haematologica. 2010. PMID: 20435622 Free PMC article.
-
The genetics of antithrombin.Thromb Res. 2018 Sep;169:23-29. doi: 10.1016/j.thromres.2018.07.008. Epub 2018 Jul 5. Thromb Res. 2018. PMID: 30005274 Review.
-
Antiangiogenic function of antithrombin is dependent on its conformational variation: implication for other serpins.Protein Pept Lett. 2013 Apr;20(4):403-11. Protein Pept Lett. 2013. PMID: 23016581 Review.
Cited by
-
The introduction of an N-glycosylation site into prochymosin greatly enhances its production and secretion by Pichia pastoris.Microb Cell Fact. 2022 Aug 30;21(1):177. doi: 10.1186/s12934-022-01904-3. Microb Cell Fact. 2022. PMID: 36042512 Free PMC article.
-
Anticoagulant SERPINs: Endogenous Regulators of Hemostasis and Thrombosis.Front Cardiovasc Med. 2022 May 3;9:878199. doi: 10.3389/fcvm.2022.878199. eCollection 2022. Front Cardiovasc Med. 2022. PMID: 35592395 Free PMC article. Review.
-
Identification and characterization of two SERPINC1 mutations causing congenital antithrombin deficiency.Thromb J. 2023 Jan 9;21(1):3. doi: 10.1186/s12959-022-00443-6. Thromb J. 2023. PMID: 36624481 Free PMC article.
-
Clinical and Molecular Characterization of Nine Novel Antithrombin Mutations.Int J Mol Sci. 2024 Mar 1;25(5):2893. doi: 10.3390/ijms25052893. Int J Mol Sci. 2024. PMID: 38474138 Free PMC article.
-
Different N-Glycosylation Sites Reduce the Activity of Recombinant DSPAα2.Curr Issues Mol Biol. 2022 Aug 31;44(9):3930-3947. doi: 10.3390/cimb44090270. Curr Issues Mol Biol. 2022. PMID: 36135182 Free PMC article.
References
-
- Lijfering W.M., Brouwer J.L.P., Veeger N.J.G.M., Bank I., Coppens M., Middeldorp S., Hamulyák K., Prins M.H., Büller H.R., Van Der Meer J. Selective testing for thrombophilia in patients with first venous thrombosis: Results from a retrospective family cohort study on absolute thrombotic risk for currently known thrombophilic defects in 2479 relatives. Blood. 2009;113:5314–5322. doi: 10.1182/blood-2008-10-184879. - DOI - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
