

Why call it developmental bias when it is just development?
- PMID: 33422150
- PMCID: PMC7796527
- DOI: 10.1186/s13062-020-00289-w
Why call it developmental bias when it is just development?
Abstract
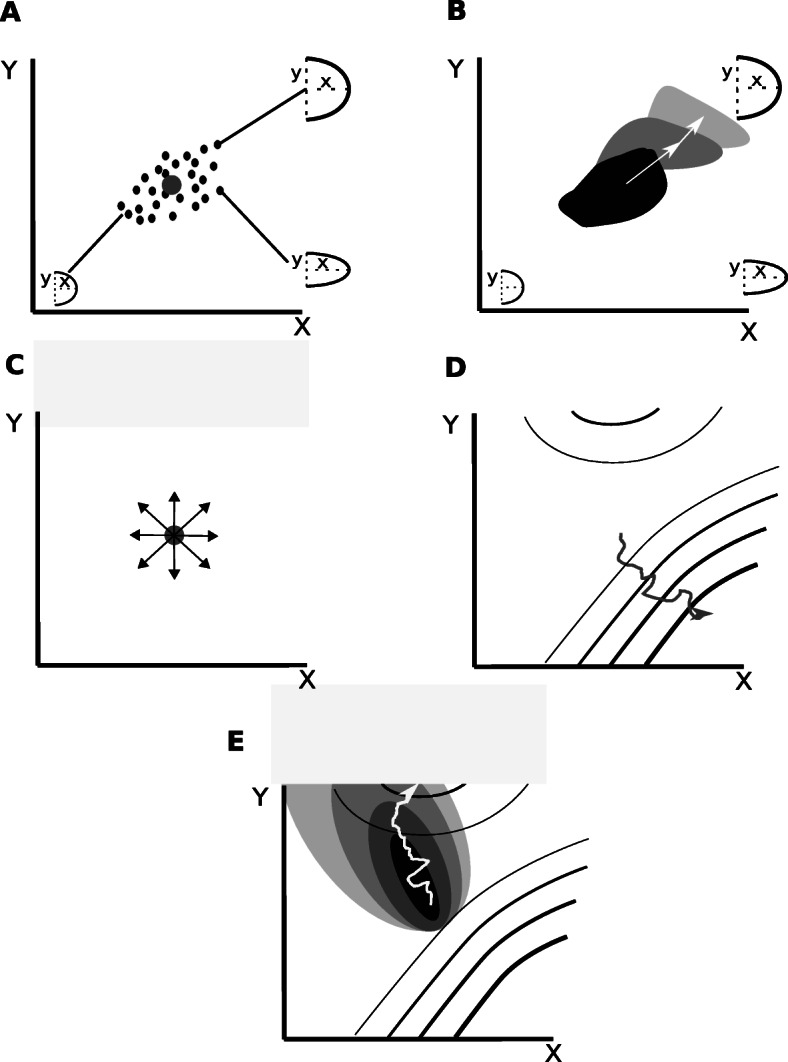
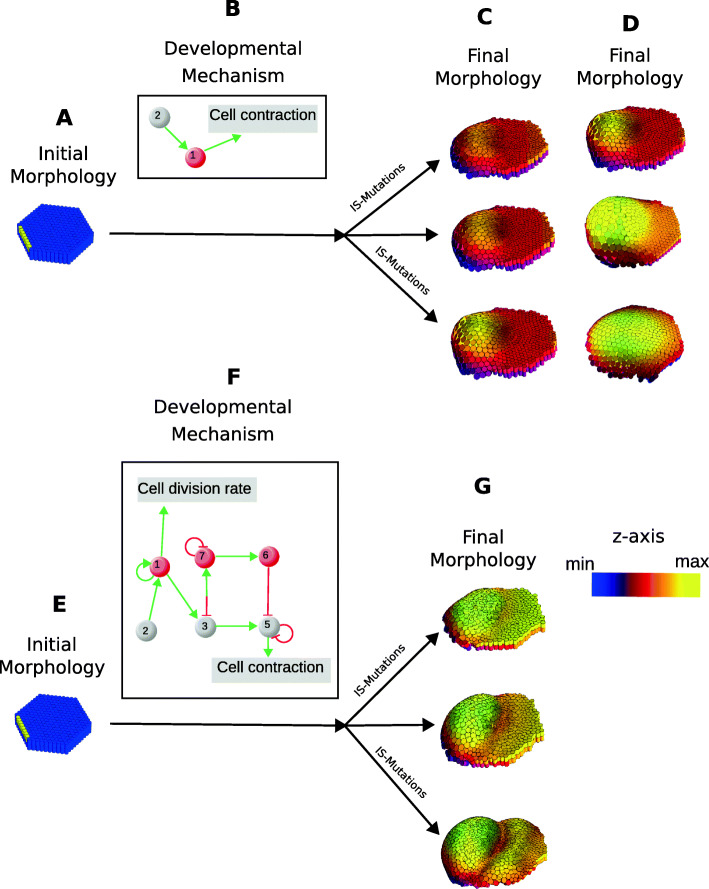
The concept of developmental constraints has been central to understand the role of development in morphological evolution. Developmental constraints are classically defined as biases imposed by development on the distribution of morphological variation.This opinion article argues that the concepts of developmental constraints and developmental biases do not accurately represent the role of development in evolution. The concept of developmental constraints was coined to oppose the view that natural selection is all-capable and to highlight the importance of development for understanding evolution. In the modern synthesis, natural selection was seen as the main factor determining the direction of morphological evolution. For that to be the case, morphological variation needs to be isotropic (i.e. equally possible in all directions). The proponents of the developmental constraint concept argued that development makes that some morphological variation is more likely than other (i.e. variation is not isotropic), and that, thus, development constraints evolution by precluding natural selection from being all-capable.This article adds to the idea that development is not compatible with the isotropic expectation by arguing that, in fact, it could not be otherwise: there is no actual reason to expect that development could lead to isotropic morphological variation. It is then argued that, since the isotropic expectation is untenable, the role of development in evolution should not be understood as a departure from such an expectation. The role of development in evolution should be described in an exclusively positive way, as the process determining which directions of morphological variation are possible, instead of negatively, as a process precluding the existence of morphological variation we have no actual reason to expect.This article discusses that this change of perspective is not a mere question of semantics: it leads to a different interpretation of the studies on developmental constraints and to a different research program in evolution and development. This program does not ask whether development constrains evolution. Instead it asks questions such as, for example, how different types of development lead to different types of morphological variation and, together with natural selection, determine the directions in which different lineages evolve.
Keywords: Development; Developmental bias; Developmental constraints; Developmental mechanisms; Morphogenesis; Morphological evolution; Variational properties.
Conflict of interest statement
The author declares that he has no competing interests.
Figures
Similar articles
-
Developmental plasticity and evolutionary explanations.Evol Dev. 2020 Jan;22(1-2):47-55. doi: 10.1111/ede.12314. Epub 2019 Sep 19. Evol Dev. 2020. PMID: 31535438
-
Does phenotypic plasticity initiate developmental bias?Evol Dev. 2020 Jan;22(1-2):56-70. doi: 10.1111/ede.12304. Epub 2019 Jul 26. Evol Dev. 2020. PMID: 31348849 Free PMC article. Review.
-
Developmental constraints versus flexibility in morphological evolution.Nature. 2002 Apr 25;416(6883):844-7. doi: 10.1038/416844a. Nature. 2002. PMID: 11976682
-
Historical and philosophical perspectives on the study of developmental bias.Evol Dev. 2020 Jan;22(1-2):7-19. doi: 10.1111/ede.12302. Epub 2019 Jul 29. Evol Dev. 2020. PMID: 31356725
-
Developmental constraints vs. variational properties: How pattern formation can help to understand evolution and development.J Exp Zool B Mol Dev Evol. 2006 Mar 15;306(2):107-25. doi: 10.1002/jez.b.21078. J Exp Zool B Mol Dev Evol. 2006. PMID: 16254986 Review.
Cited by
-
The logic of monsters: development and morphological diversity in stem-cell-based embryo models.Interface Focus. 2024 Oct 25;14(5):20240023. doi: 10.1098/rsfs.2024.0023. eCollection 2024 Oct 11. Interface Focus. 2024. PMID: 39464644 Free PMC article. Review.
-
A Grand Challenge in Development and Evodevo: Quantifying the Role of Development in Evolution.Front Plant Sci. 2022 Jan 11;12:752344. doi: 10.3389/fpls.2021.752344. eCollection 2021. Front Plant Sci. 2022. PMID: 35087543 Free PMC article. No abstract available.
-
Predicting phenotype transition probabilities via conditional algorithmic probability approximations.J R Soc Interface. 2022 Dec;19(197):20220694. doi: 10.1098/rsif.2022.0694. Epub 2022 Dec 14. J R Soc Interface. 2022. PMID: 36514888 Free PMC article.
-
The structure of an ancient genotype-phenotype map shaped the functional evolution of a protein family.Nat Ecol Evol. 2025 Jul 25. doi: 10.1038/s41559-025-02777-6. Online ahead of print. Nat Ecol Evol. 2025. PMID: 40715711
-
The legacy and evolvability of Pere Alberch's ideas.Interface Focus. 2024 Oct 25;14(5):20240011. doi: 10.1098/rsfs.2024.0011. eCollection 2024 Oct 11. Interface Focus. 2024. PMID: 39464645 Free PMC article.
References
-
- Darwin C. On the Origin of Species. London: John Murray; 1859.
-
- Waddington CH. The strategy of the genes. London: George Allen Unwin Ltd; 1957.
-
- Alberch P. Ontogenesis and morphological diversification. Amer Zool. 1980;20:653–667. doi: 10.1093/icb/20.4.653. - DOI
-
- Wake DB, Roth G, Wake MH. On the problem of stasis in organismal evolution. J Theor Biol. 1983;101:211–224. doi: 10.1016/0022-5193(83)90335-1. - DOI
-
- Maynard Smith J, Burian R, Kauffman SA, Alberch P, et al. Developmental constrains and evolution. Q Rev Biol. 1985;60:265–287. doi: 10.1086/414425. - DOI
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
