

Uncovering the Role of p38 Family Members in Adipose Tissue Physiology
- PMID: 33424765
- PMCID: PMC7786386
- DOI: 10.3389/fendo.2020.572089
Uncovering the Role of p38 Family Members in Adipose Tissue Physiology
Abstract
The complex functions of adipose tissue have been a focus of research interest over the past twenty years. Adipose tissue is not only the main energy storage depot, but also one of the largest endocrine organs in the body and carries out crucial metabolic functions. Moreover, brown and beige adipose depots are major sites of energy expenditure through the activation of adaptive, non-shivering thermogenesis. In recent years, numerous signaling molecules and pathways have emerged as critical regulators of adipose tissue, in both homeostasis and obesity-related disease. Among the best characterized are members of the p38 kinase family. The activity of these kinases has emerged as a key contributor to the biology of the white and brown adipose tissues, and their modulation could provide new therapeutic approaches against obesity. Here, we give an overview of the roles of the distinct p38 family members in adipose tissue, focusing on their actions in adipogenesis, thermogenic activity, and secretory function.
Keywords: Adipose; Brown; P38; kinase; p38 mitogen-activated protein kinase(s); signaling.
Copyright © 2020 Leiva, Matesanz, Pulgarín-Alfaro, Nikolic and Sabio.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures
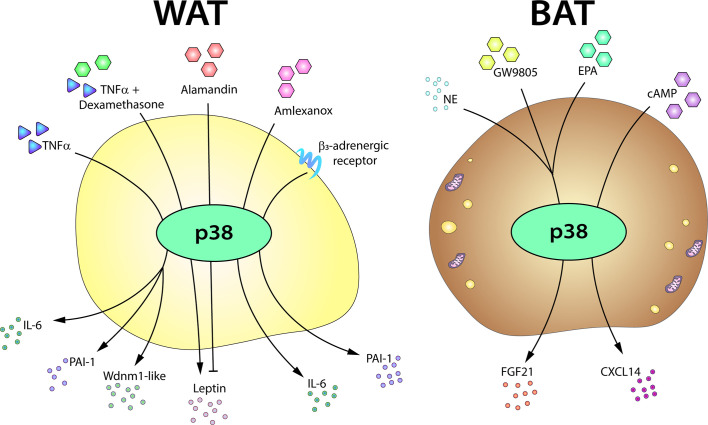
Similar articles
-
GDF5 Promotes White Adipose Tissue Thermogenesis via p38 MAPK Signaling Pathway.DNA Cell Biol. 2019 Nov;38(11):1303-1312. doi: 10.1089/dna.2019.4724. Epub 2019 Sep 25. DNA Cell Biol. 2019. PMID: 31553232
-
White, brown, beige/brite: different adipose cells for different functions?Endocrinology. 2013 Sep;154(9):2992-3000. doi: 10.1210/en.2013-1403. Epub 2013 Jun 19. Endocrinology. 2013. PMID: 23782940 Review.
-
The role of autophagy in brown and beige adipose tissue plasticity.J Physiol Biochem. 2020 May;76(2):213-226. doi: 10.1007/s13105-019-00708-1. Epub 2019 Dec 6. J Physiol Biochem. 2020. PMID: 31811543 Review.
-
Browning and thermogenic programing of adipose tissue.Best Pract Res Clin Endocrinol Metab. 2016 Aug;30(4):479-485. doi: 10.1016/j.beem.2016.09.003. Epub 2016 Sep 12. Best Pract Res Clin Endocrinol Metab. 2016. PMID: 27697209 Review.
-
Prior exercise training improves cold tolerance independent of indices associated with non-shivering thermogenesis.J Physiol. 2018 Sep;596(18):4375-4391. doi: 10.1113/JP276228. Epub 2018 Aug 14. J Physiol. 2018. PMID: 30109697 Free PMC article.
Cited by
-
Echinacoside Induces UCP1- and ATP-Dependent Thermogenesis in Beige Adipocytes via the Activation of Dopaminergic Receptors.J Microbiol Biotechnol. 2023 Oct 28;33(10):1268-1280. doi: 10.4014/jmb.2306.06041. Epub 2023 Jul 17. J Microbiol Biotechnol. 2023. PMID: 37463854 Free PMC article.
-
Adipose tissue as target of environmental toxicants: focus on mitochondrial dysfunction and oxidative inflammation in metabolic dysfunction-associated steatotic liver disease.Mol Cell Biochem. 2025 May;480(5):2863-2879. doi: 10.1007/s11010-024-05165-z. Epub 2024 Dec 20. Mol Cell Biochem. 2025. PMID: 39704874 Free PMC article. Review.
-
IL-27 increases energy storage in white adipocytes by enhancing glucose uptake and fatty acid esterification.Adipocyte. 2023 Dec;12(1):2276346. doi: 10.1080/21623945.2023.2276346. Epub 2023 Nov 10. Adipocyte. 2023. PMID: 37948192 Free PMC article.
-
A nexus of lipid and O-Glcnac metabolism in physiology and disease.Front Endocrinol (Lausanne). 2022 Aug 30;13:943576. doi: 10.3389/fendo.2022.943576. eCollection 2022. Front Endocrinol (Lausanne). 2022. PMID: 36111295 Free PMC article. Review.
-
Ginsenoside CK Inhibits the Early Stage of Adipogenesis via the AMPK, MAPK, and AKT Signaling Pathways.Antioxidants (Basel). 2022 Sep 23;11(10):1890. doi: 10.3390/antiox11101890. Antioxidants (Basel). 2022. PMID: 36290613 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous