

Cardinium Localization During Its Parasitoid Wasp Host's Development Provides Insights Into Cytoplasmic Incompatibility
- PMID: 33424808
- PMCID: PMC7793848
- DOI: 10.3389/fmicb.2020.606399
Cardinium Localization During Its Parasitoid Wasp Host's Development Provides Insights Into Cytoplasmic Incompatibility
Abstract
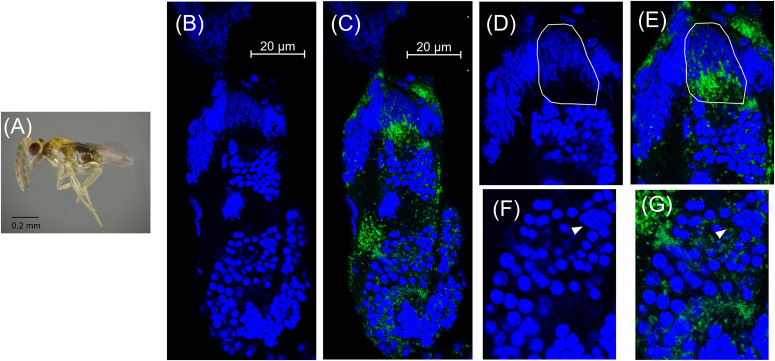
Arthropods harbor heritable intracellular symbionts that may manipulate host reproduction to favor symbiont transmission. In cytoplasmic incompatibility (CI), the symbiont sabotages the reproduction of infected males such that high levels of offspring mortality result when they mate with uninfected females. In crosses with infected males and infected females, however (the "rescue" cross), normal numbers of offspring are produced. A common CI-inducing symbiont, Cardinium hertigii, causes variable levels of CI mortality in the parasitoid wasp, Encarsia suzannae. Previous work correlated CI-induced mortality with male development time in this system, although the timing of Cardinium CI-induction and the relationship between development time and CI mortality was not well understood. Here, using a combination of crosses, manipulation of development time, and fluorescence microscopy, we identify the localization and the timing of the CI-induction step in the Cardinium-E. suzannae system. Antibiotic treatment of adult Cardinium-infected males did not reduce the mortality associated with the CI phenotype, suggesting that CI-alteration occurs prior to adulthood. Our results suggest that the alteration step occurs during the pupal period, and is limited by the duration of pupal development: 1) Encarsia produces most sperm prior to adulthood, 2) FISH localization of Cardinium in testes showed an association with sperm nuclei throughout spermatogenesis but not with mature sperm, and 3) two methods of prolonging the pupal period (cool temperatures and the juvenile hormone analog methoprene) both caused greater CI mortality, suggesting the degree of alteration is limited by the duration of the pupal stage. Based on these results, we compare two models for potential mechanisms of Cardinium sperm modification in the context of what is known about analogous mechanisms of Wolbachia, a more extensively studied CI-inducing symbiont.
Keywords: cardinium; cytoplasmic incompatibility; encarsia; parasitoid; spermatogenesis; symbiosis; wolbachia.
Copyright © 2020 Doremus, Stouthamer, Kelly, Schmitz-Esser and Hunter.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures






References
Associated data
LinkOut - more resources
Full Text Sources
Other Literature Sources