

Embryo-Endosperm Interaction and Its Agronomic Relevance to Rice Quality
- PMID: 33424883
- PMCID: PMC7793959
- DOI: 10.3389/fpls.2020.587641
Embryo-Endosperm Interaction and Its Agronomic Relevance to Rice Quality
Abstract
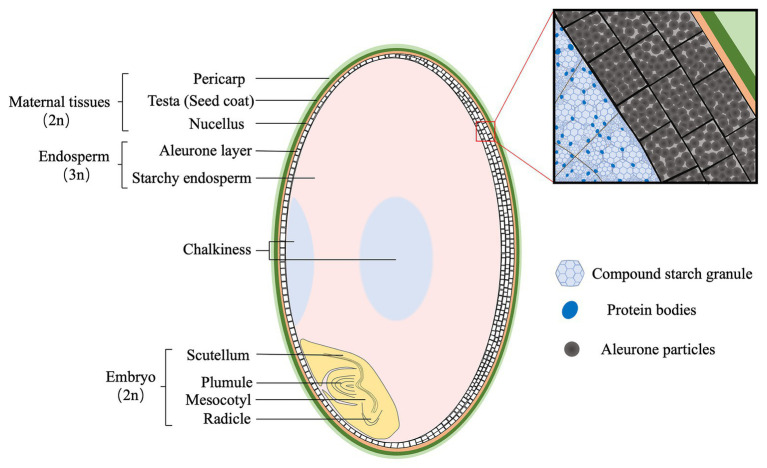
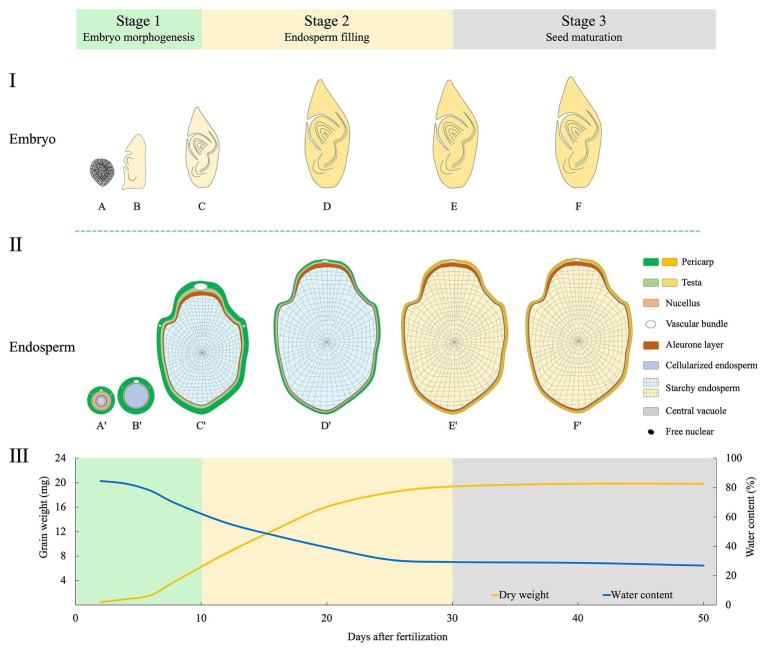
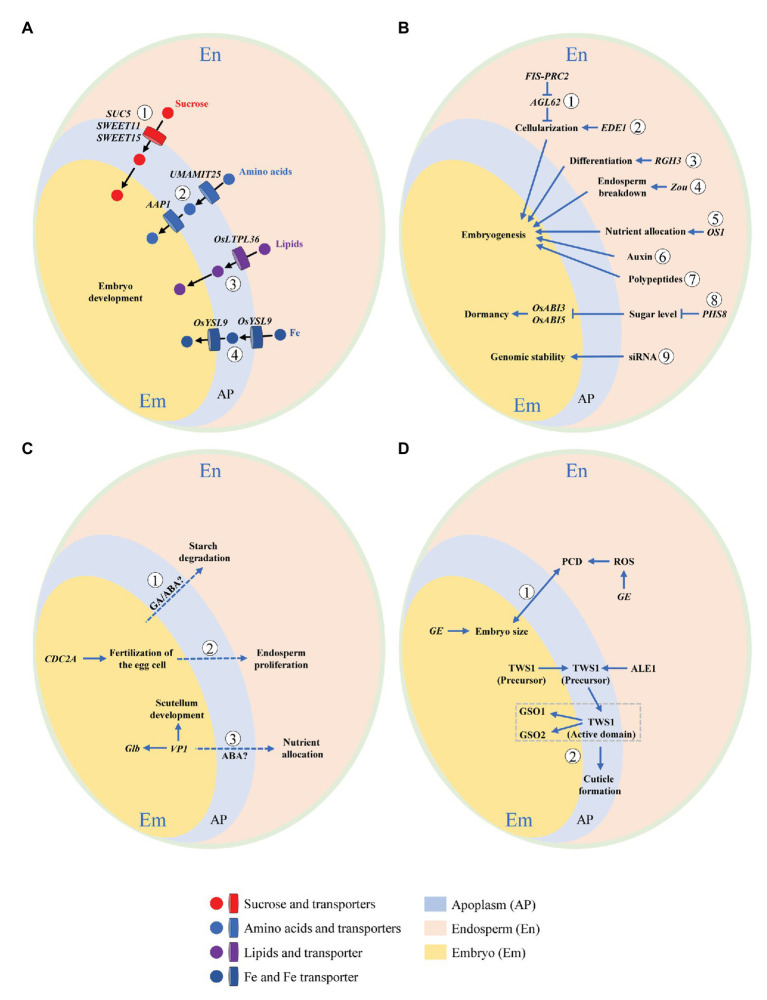
Embryo-endosperm interaction is the dominant process controlling grain filling, thus being crucial for yield and quality formation of the three most important cereals worldwide, rice, wheat, and maize. Fundamental science of functional genomics has uncovered several key genetic programs for embryo and endosperm development, but the interaction or communication between the two tissues is largely elusive. Further, the significance of this interaction for grain filling remains open. This review starts with the morphological and developmental aspects of rice grain, providing a spatial and temporal context. Then, it offers a comprehensive and integrative view of this intercompartmental interaction, focusing on (i) apoplastic nutrient flow from endosperm to the developing embryo, (ii) dependence of embryo development on endosperm, (iii) regulation of endosperm development by embryo, and (iv) bidirectional dialogues between embryo and endosperm. From perspective of embryo-endosperm interaction, the mechanisms underlying the complex quality traits are explored, with grain chalkiness as an example. The review ends with three open questions with scientific and agronomic importance that should be addressed in the future. Notably, current knowledge and future prospects of this hot research topic are reviewed from a viewpoint of crop physiology, which should be helpful for bridging the knowledge gap between the fundamental plant sciences and the practical technologies.
Keywords: embryo; endosperm; grain quality; interaction; rice; seed development.
Copyright © 2020 An, Tao, Chen, He, Xiao, Li, Ding and Liu.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures

References
-
- Bao J. S. (2019). “Rice milling quality” in Rice. 4th Edn. (Duxford: Elsevier; ), 339–369.
-
- Belmonte M. F., Kirkbride R. C., Stone S. L., Pelletier J. M., Bui A. Q., Yeung E. C., et al. . (2013). Comprehensive developmental profiles of gene activity in regions and subregions of the Arabidopsis seed. Proc. Natl. Acad. Sci. U. S. A. 110, E435–E444. 10.1073/pnas.1222061110, PMID: - DOI - PMC - PubMed
Publication types
LinkOut - more resources
Full Text Sources
Miscellaneous