

Disrupting the Homeostasis of High Mobility Group Protein Promotes the Systemic Movement of Bamboo mosaic virus
- PMID: 33424893
- PMCID: PMC7793662
- DOI: 10.3389/fpls.2020.597665
Disrupting the Homeostasis of High Mobility Group Protein Promotes the Systemic Movement of Bamboo mosaic virus
Abstract
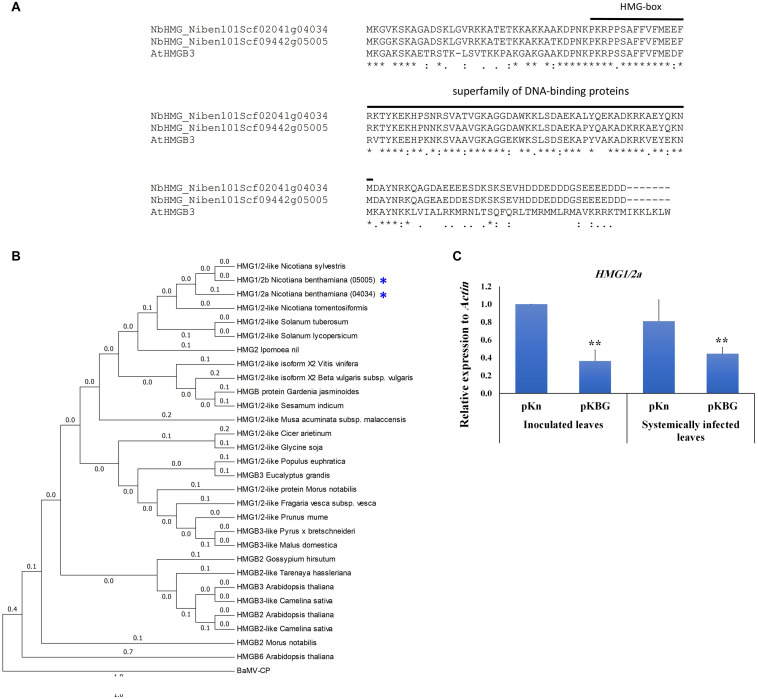
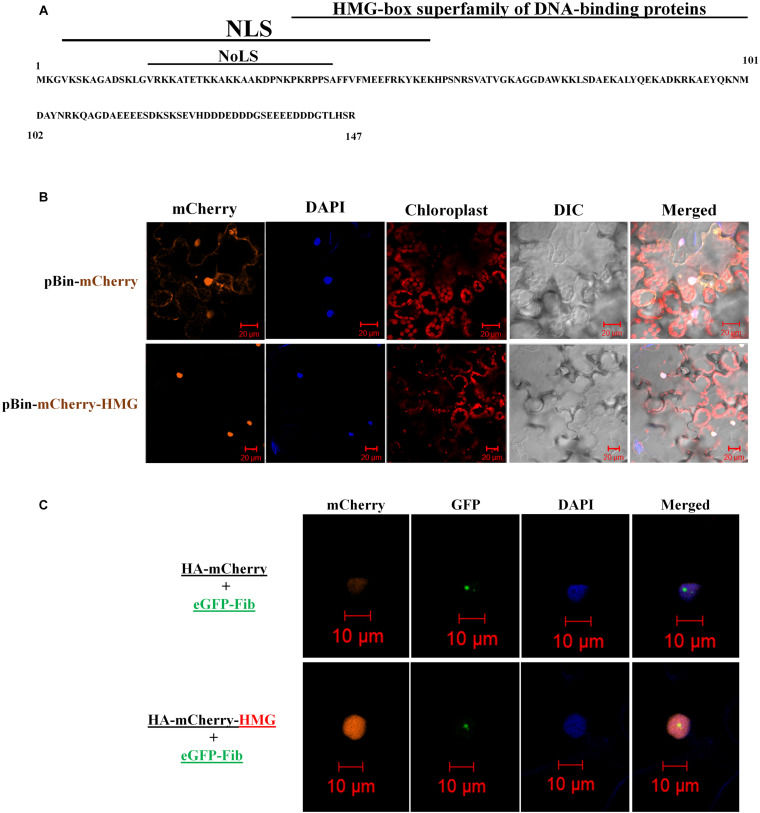
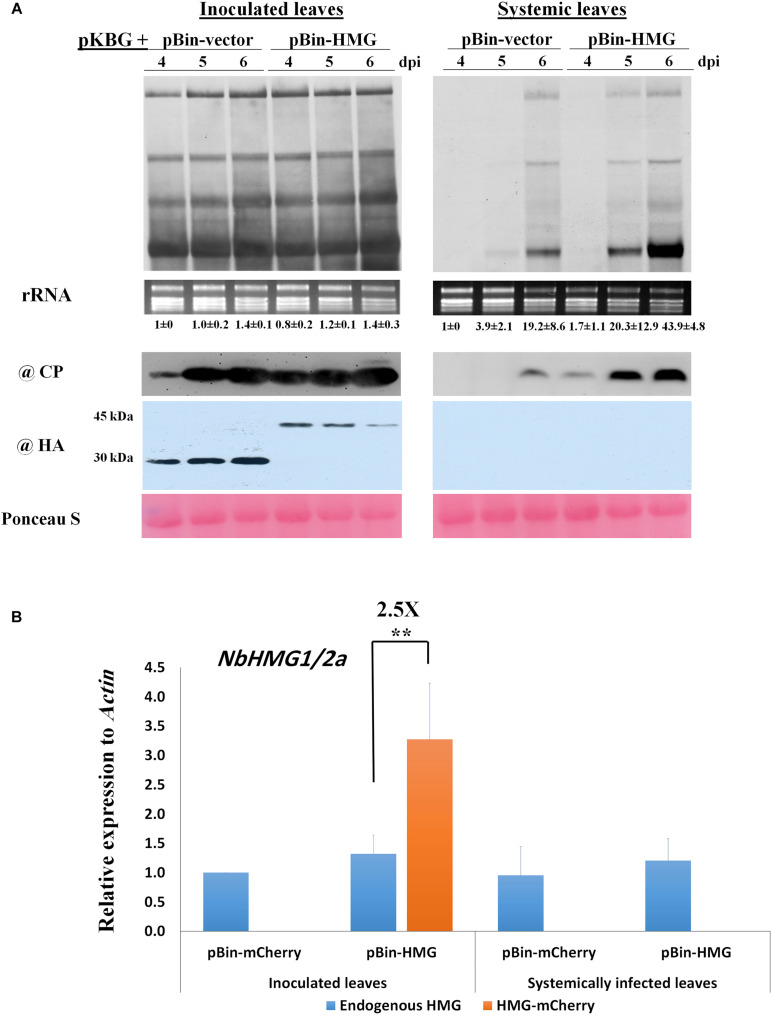
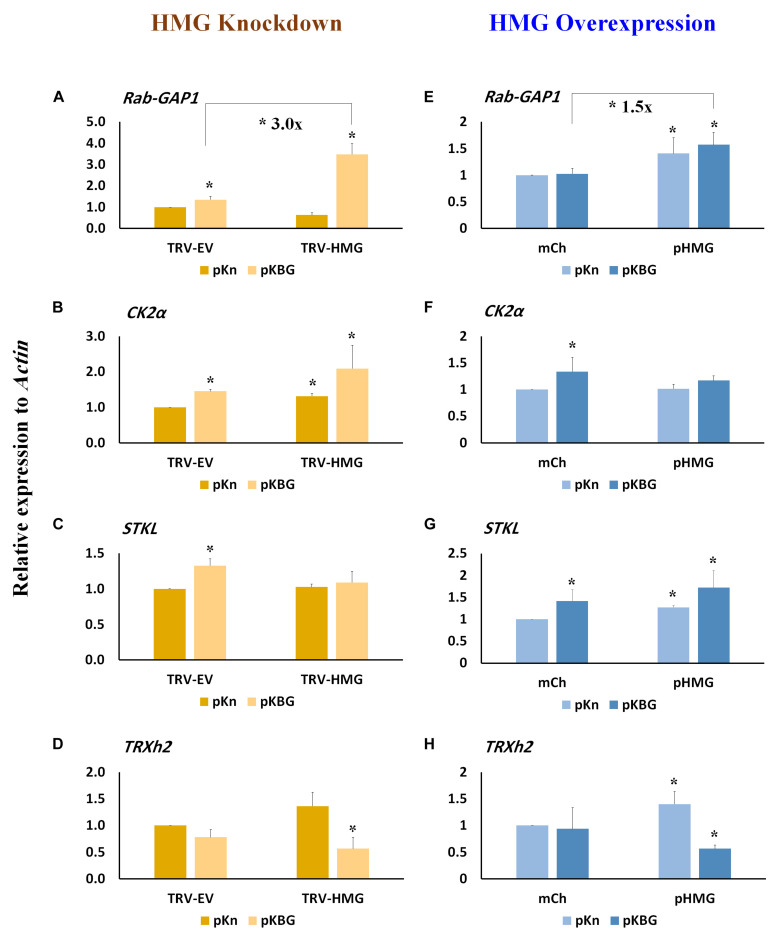
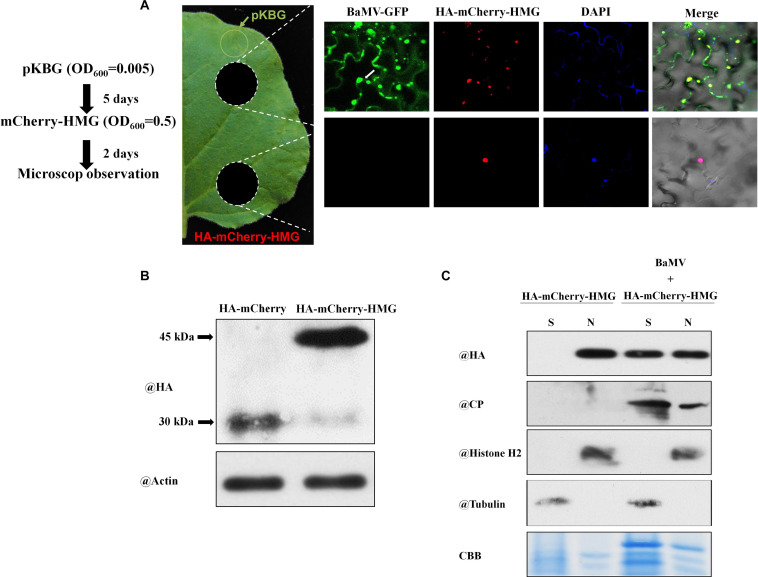
Viruses hijack various organelles and machineries for their replication and movement. Ever more lines of evidence indicate that specific nuclear factors are involved in systemic trafficking of several viruses. However, how such factors regulate viral systemic movement remains unclear. Here, we identify a novel role for Nicotiana benthamiana high mobility group nucleoprotein (NbHMG1/2a) in virus movement. Although infection of N. benthamiana with Bamboo mosaic virus (BaMV) decreased NbHMG1/2a expression levels, nuclear-localized NbHMG1/2a protein was shuttled out of the nucleus into cytoplasm upon BaMV infection. NbHMG1/2a knockdown or even overexpression did not affect BaMV accumulation in inoculated leaves, but it did enhance systemic movement of the virus. Interestingly, the positive regulator Rap-GTPase activation protein 1 was highly upregulated upon infection with BaMV, whereas the negative regulator thioredoxin h protein was greatly reduced, no matter if NbHMG1a/2a was silenced or overexpressed. Our findings indicate that NbHMG1/2a may have a role in plant defense responses. Once its homeostasis is disrupted, expression of relevant host factors may be perturbed that, in turn, facilitates BaMV systemic movement.
Keywords: Bamboo mosaic virus; high mobility group proteins; nucleoprotein; plant-virus interaction; virus movement.
Copyright © 2020 Alazem, He, Chang, Cheng and Lin.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures

References
LinkOut - more resources
Full Text Sources
Research Materials