

Overcoming Energetic Barriers in Acetogenic C1 Conversion
- PMID: 33425882
- PMCID: PMC7793690
- DOI: 10.3389/fbioe.2020.621166
Overcoming Energetic Barriers in Acetogenic C1 Conversion
Abstract
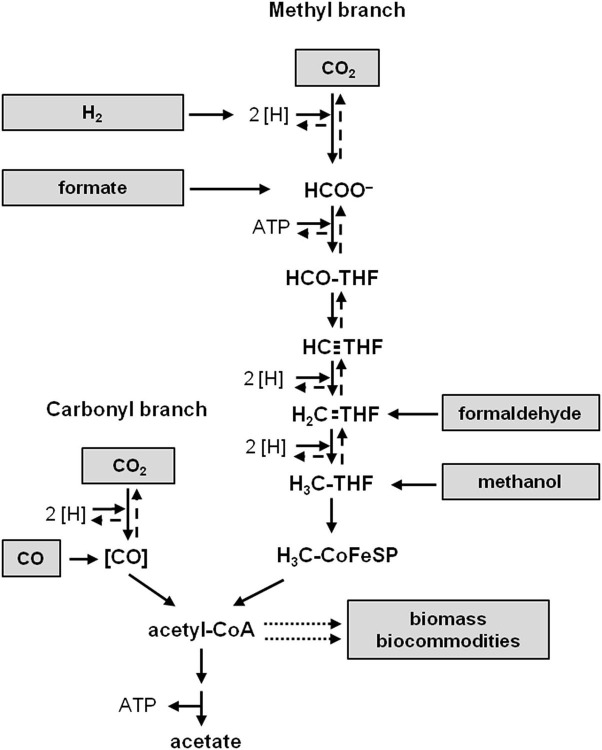
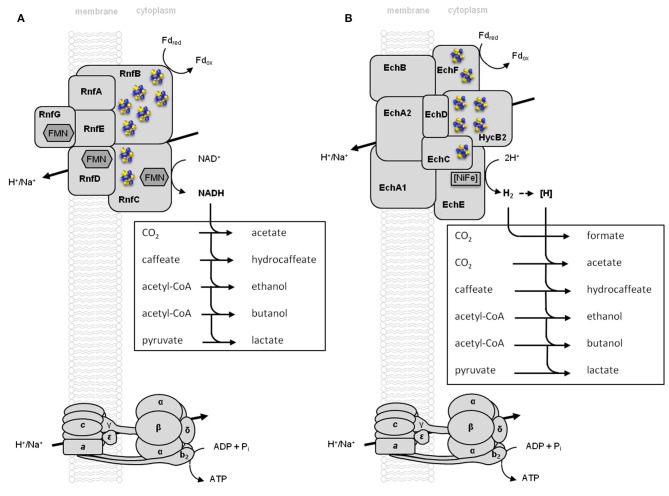
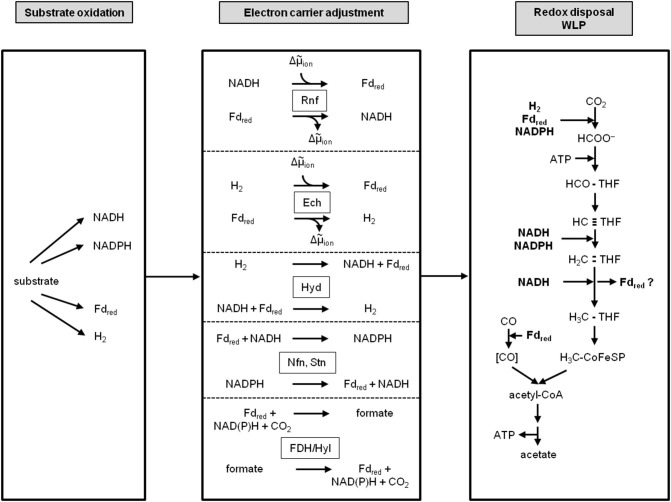
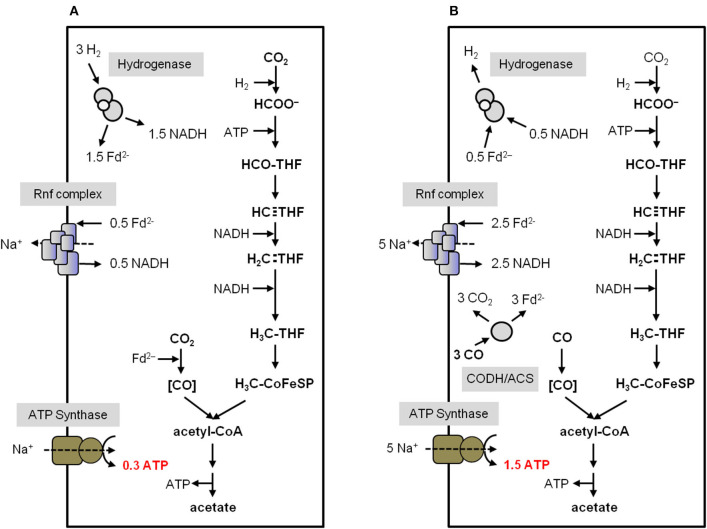
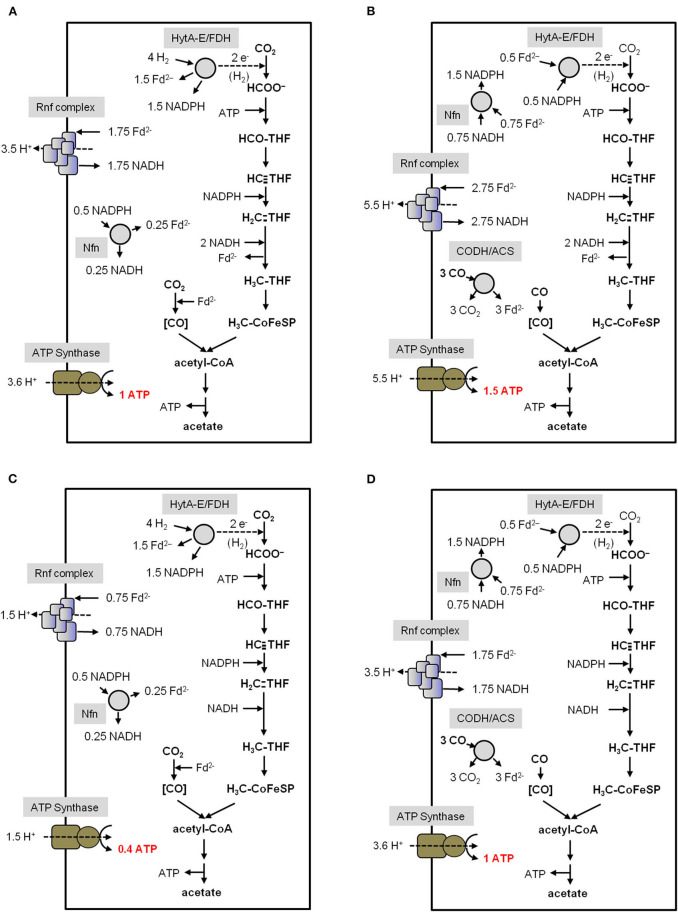
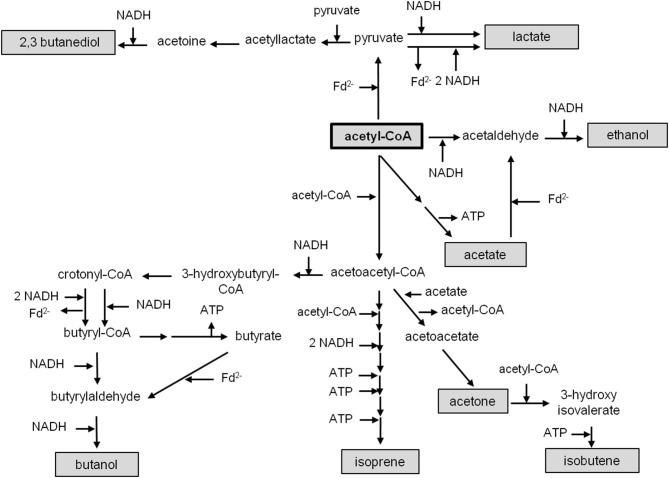
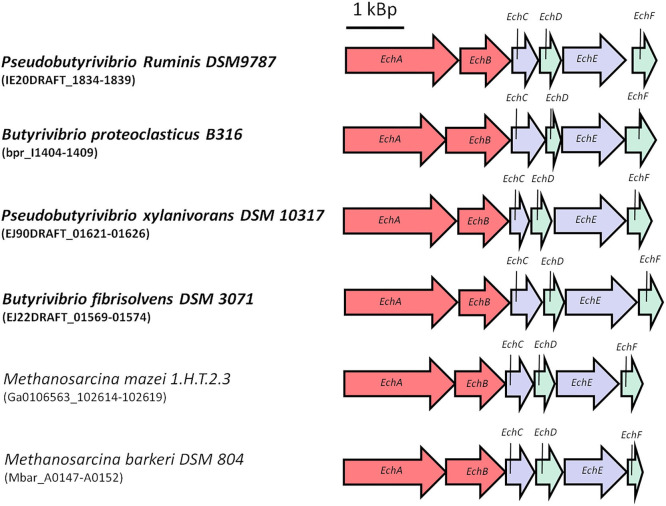
Currently one of the biggest challenges for society is to combat global warming. A solution to this global threat is the implementation of a CO2-based bioeconomy and a H2-based bioenergy economy. Anaerobic lithotrophic bacteria such as the acetogenic bacteria are key players in the global carbon and H2 cycle and thus prime candidates as driving forces in a H2- and CO2-bioeconomy. Naturally, they convert two molecules of CO2 via the Wood-Ljungdahl pathway (WLP) to one molecule of acetyl-CoA which can be converted to different C2-products (acetate or ethanol) or elongated to C4 (butyrate) or C5-products (caproate). Since there is no net ATP generation from acetate formation, an electron-transport phosphorylation (ETP) module is hooked up to the WLP. ETP provides the cell with additional ATP, but the ATP gain is very low, only a fraction of an ATP per mol of acetate. Since acetogens live at the thermodynamic edge of life, metabolic engineering to obtain high-value products is currently limited by the low energy status of the cells that allows for the production of only a few compounds with rather low specificity. To set the stage for acetogens as production platforms for a wide range of bioproducts from CO2, the energetic barriers have to be overcome. This review summarizes the pathway, the energetics of the pathway and describes ways to overcome energetic barriers in acetogenic C1 conversion.
Keywords: biofuels; biohydrogen; carbon capture; electron transport; hydrogen storage.
Copyright © 2020 Katsyv and Müller.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures
References
-
- Abken H. J., Tietze M., Brodersen J., Baumer S., Beifuss U., Deppenmeier U. (1998). Isolation and characterization of methanophenazine and function of phenazines in membrane-bound electron transport of Methanosarcina mazei Gö1. J. Bacteriol. 180, 2027–2032. 10.1128/JB.180.8.2027-2032.1998 - DOI - PMC - PubMed
-
- Abrini J., Naveau H., Nyns E. J. (1994). Clostridium autoethanogenum, sp. nov., an anaerobic bacterium that produces ethanol from carbon monoxide. Arch. Microbiol. 161, 345–351. 10.1007/BF00303591 - DOI
-
- Ajanovic A. (2011). Biofuels versus food production: does biofuels production increase food prices? Energy 36, 2070–2076. 10.1016/j.energy.2010.05.019 - DOI
Publication types
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous