

Proteostasis of Islet Amyloid Polypeptide: A Molecular Perspective of Risk Factors and Protective Strategies for Type II Diabetes
- PMID: 33427465
- PMCID: PMC10317076
- DOI: 10.1021/acs.chemrev.0c00981
Proteostasis of Islet Amyloid Polypeptide: A Molecular Perspective of Risk Factors and Protective Strategies for Type II Diabetes
Abstract
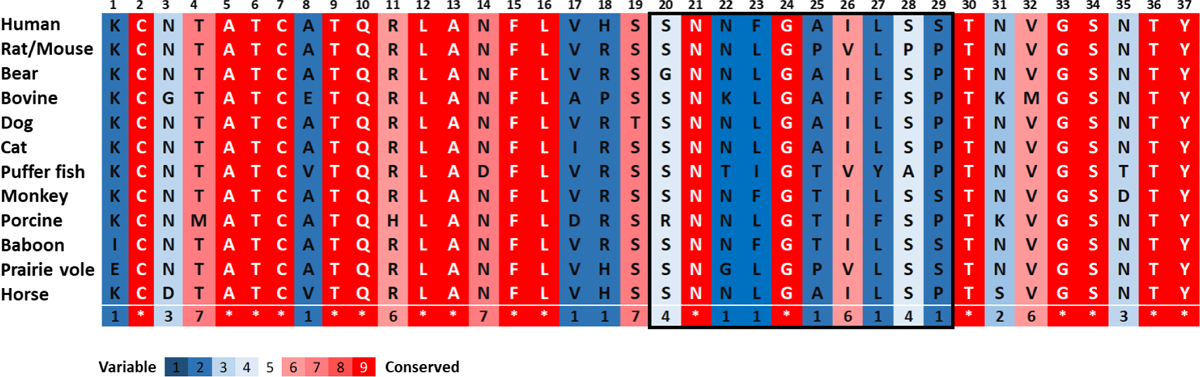
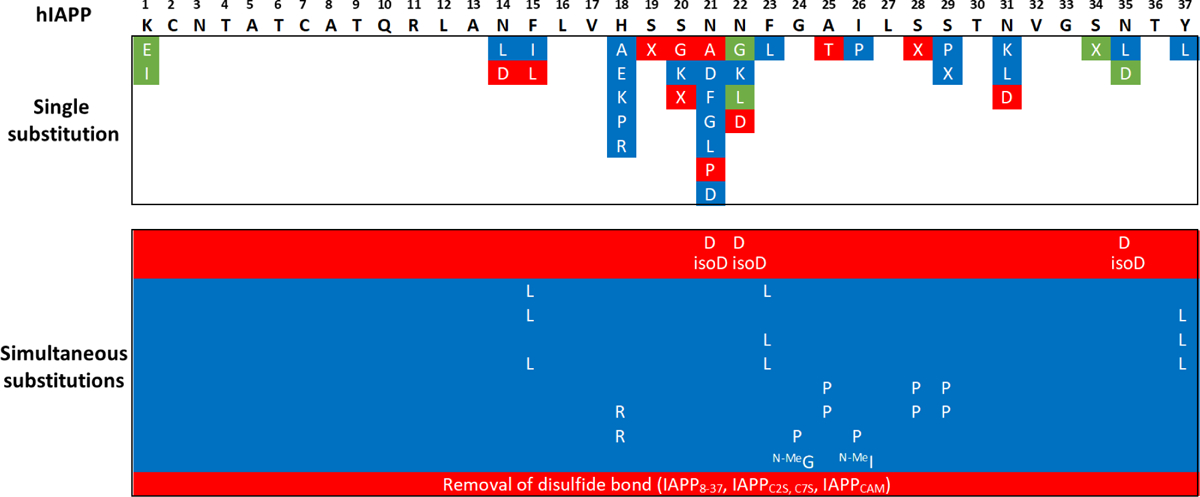
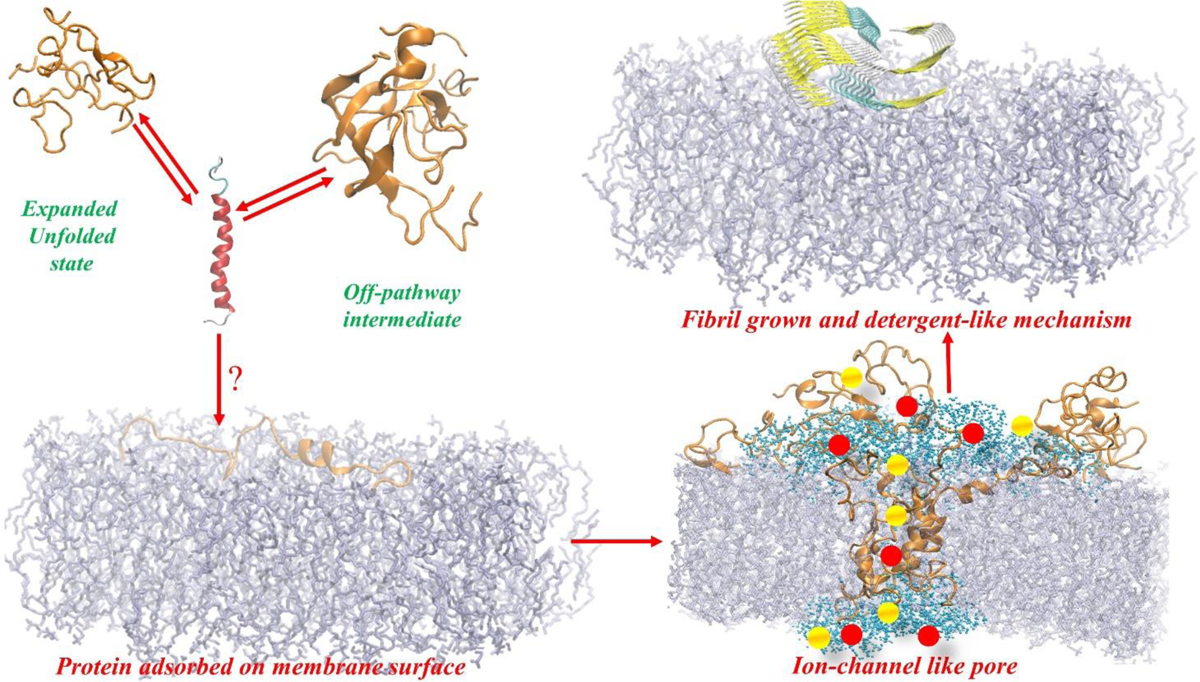
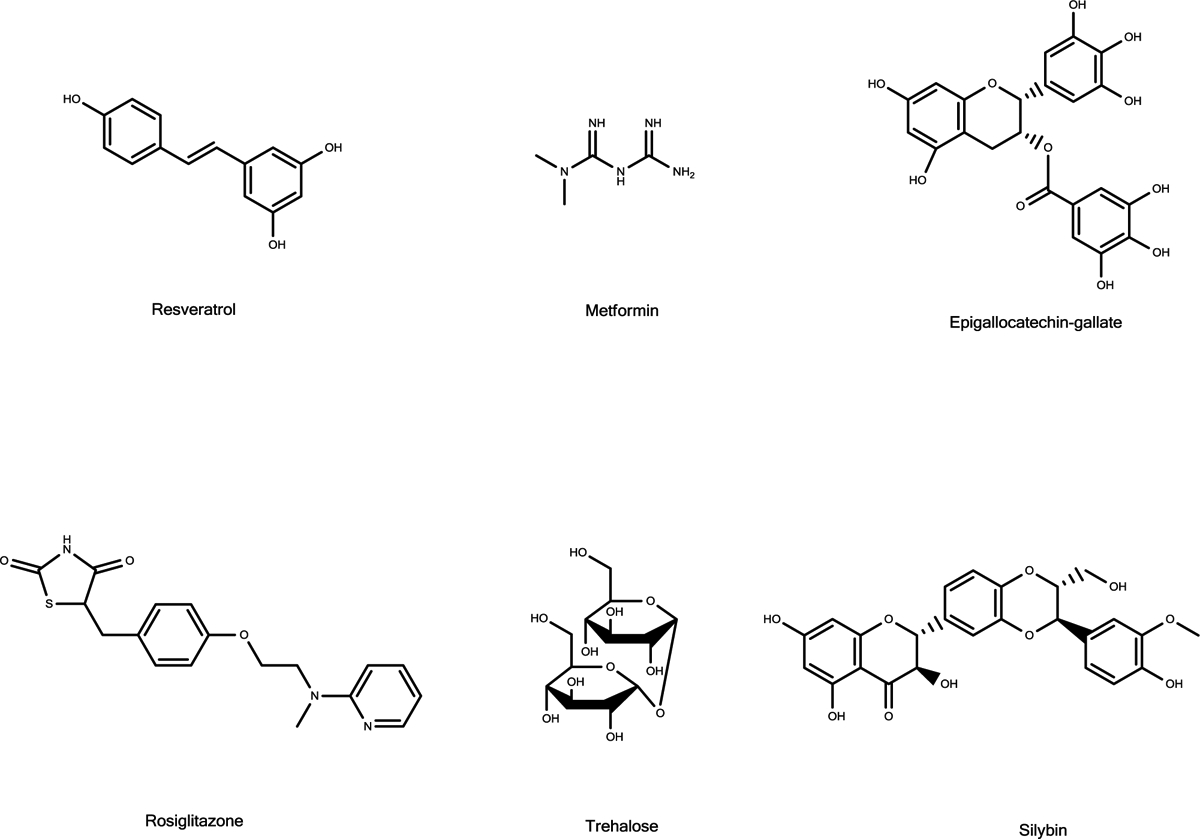
The possible link between hIAPP accumulation and β-cell death in diabetic patients has inspired numerous studies focusing on amyloid structures and aggregation pathways of this hormone. Recent studies have reported on the importance of early oligomeric intermediates, the many roles of their interactions with lipid membrane, pH, insulin, and zinc on the mechanism of aggregation of hIAPP. The challenges posed by the transient nature of amyloid oligomers, their structural heterogeneity, and the complex nature of their interaction with lipid membranes have resulted in the development of a wide range of biophysical and chemical approaches to characterize the aggregation process. While the cellular processes and factors activating hIAPP-mediated cytotoxicity are still not clear, it has recently been suggested that its impaired turnover and cellular processing by proteasome and autophagy may contribute significantly toward toxic hIAPP accumulation and, eventually, β-cell death. Therefore, studies focusing on the restoration of hIAPP proteostasis may represent a promising arena for the design of effective therapies. In this review we discuss the current knowledge of the structures and pathology associated with hIAPP self-assembly and point out the opportunities for therapy that a detailed biochemical, biophysical, and cellular understanding of its aggregation may unveil.
Figures




References
-
- Schwartz P New patho-anatomic observations on amyloidosis in the aged. Fluorescence microscopic investigations. In Amyloidosis; Mandema E, Ruinen L, Scholten JH, Cohen AS, Eds.; Excerpta Medica: Amsterdam, 1968; pp 400–15.
-
- Westermark P Islet Amyloid Polypeptide and Amyloid in the Islets of Langerhans; Leslie RDG, Robbins D, Eds.; Cambridge University Press: Cambridge, UK, 1995.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
