

Prognostic Value and Function of KLF5 in Papillary Thyroid Cancer
- PMID: 33430300
- PMCID: PMC7825749
- DOI: 10.3390/cancers13020185
Prognostic Value and Function of KLF5 in Papillary Thyroid Cancer
Abstract
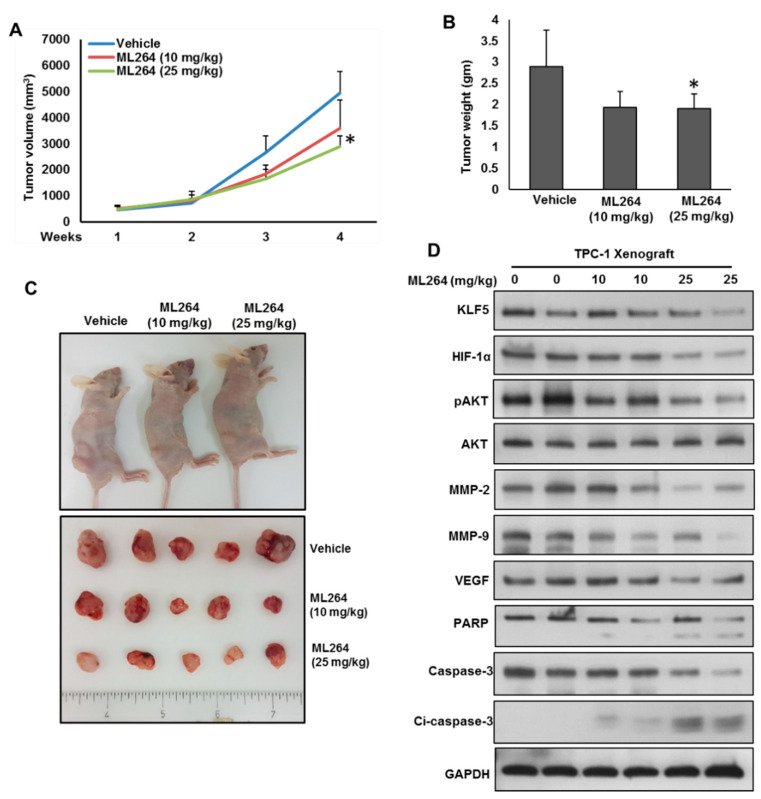
The Krüppel-like factor 5 (KLF5), a zinc-finger transcriptional factor, is highly expressed in several solid tumors, but its role in PTC remains unclear. We investigated the expression of KLF5 protein in a large cohort of PTC patient samples and explored its functional role and mechanism in PTC cell lines in vitro and in vivo. KLF5 overexpression was observed in 65.1% of all PTC cases and it was significantly associated with aggressive clinico-pathological parameters and poor outcome. Given the significant association between KLF5 and HIF-1α overexpression in PTC patients, we investigated the functional correlation between KLF5 and HIF-1α in PTC cells. Indeed, the analysis revealed the co-immunoprecipitation of KLF5 with HIF-1α in PTC cells. We also identified KLF5-binding sites in the HIF-1α promoter that specifically bound to KLF5 protein. Mechanistically, KLF5 promoted PTC cell growth, invasion, migration, and angiogenesis, while KLF5 downregulation via specific inhibitor or siRNA reverses its action in vitro. Importantly, the silencing of KLF5 decreases the self-renewal ability of spheroids generated from PTC cells. In addition, the depletion of KLF5 reduces PTC xenograft growth in vivo. These findings suggest KLF5 can be a possible new molecular therapeutic target for a subset of PTC.
Keywords: HIF-1α; KLF5; apoptosis; invasion; papillary thyroid cancer; stemness.
Conflict of interest statement
The authors declare no conflict of interest.
Figures





Similar articles
-
KLF5 promotes hypoxia-induced survival and inhibits apoptosis in non-small cell lung cancer cells via HIF-1α.Int J Oncol. 2014 Oct;45(4):1507-14. doi: 10.3892/ijo.2014.2544. Epub 2014 Jul 17. Int J Oncol. 2014. PMID: 25051115
-
Crocin inhibits the migration, invasion, and epithelial-mesenchymal transition of gastric cancer cells via miR-320/KLF5/HIF-1α signaling.J Cell Physiol. 2019 Aug;234(10):17876-17885. doi: 10.1002/jcp.28418. Epub 2019 Mar 9. J Cell Physiol. 2019. PMID: 30851060
-
Up-regulation of Krüppel-like factor 5 in pancreatic cancer is promoted by interleukin-1beta signaling and hypoxia-inducible factor-1alpha.Mol Cancer Res. 2009 Aug;7(8):1390-8. doi: 10.1158/1541-7786.MCR-08-0525. Epub 2009 Aug 11. Mol Cancer Res. 2009. PMID: 19671674
-
Krupple-Like Factor 5 is a Potential Therapeutic Target and Prognostic Marker in Epithelial Ovarian Cancer.Front Pharmacol. 2020 Dec 3;11:598880. doi: 10.3389/fphar.2020.598880. eCollection 2020. Front Pharmacol. 2020. PMID: 33424607 Free PMC article.
-
KLF5 mediates vascular remodeling via HIF-1α in hypoxic pulmonary hypertension.Am J Physiol Lung Cell Mol Physiol. 2016 Feb 15;310(4):L299-310. doi: 10.1152/ajplung.00189.2015. Epub 2015 Dec 23. Am J Physiol Lung Cell Mol Physiol. 2016. PMID: 26702149
Cited by
-
Reprogramming of Thyroid Cancer Metabolism: from Mechanism to Therapeutic Strategy.Mol Cancer. 2025 Mar 11;24(1):74. doi: 10.1186/s12943-025-02263-4. Mol Cancer. 2025. PMID: 40069775 Free PMC article. Review.
-
Inhibitory effect of the novel tyrosine kinase inhibitor DCC-2036 on triple-negative breast cancer stem cells through AXL-KLF5 positive feedback loop.Cell Death Dis. 2022 Aug 30;13(8):749. doi: 10.1038/s41419-022-05185-x. Cell Death Dis. 2022. PMID: 36042208 Free PMC article.
-
Krüppel-like factors in tumors: Key regulators and therapeutic avenues.Front Oncol. 2023 Jan 25;13:1080720. doi: 10.3389/fonc.2023.1080720. eCollection 2023. Front Oncol. 2023. PMID: 36761967 Free PMC article. Review.
References
-
- Howlader N., Noone A., Krapcho M., Neyman N., Aminou R., Altekruse S., Kosary C., Ruhl J., Tatalovich Z., Cho H. SEER Cancer Statistics Review, 1975–2009 (Vintage 2009 Populations) National Cancer Institute; Bethesda, MD, USA: 2012.
-
- Mori A., Moser C., Lang S.A., Hackl C., Gottfried E., Kreutz M., Schlitt H.J., Geissler E.K., Stoeltzing O. Up-regulation of Krüppel-like factor 5 in pancreatic cancer is promoted by interleukin-1β signaling and hypoxia-inducible factor-1α. Mol. Cancer Res. 2009;7:1390–1398. doi: 10.1158/1541-7786.MCR-08-0525. - DOI - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
