

Cold-Shock Domains-Abundance, Structure, Properties, and Nucleic-Acid Binding
- PMID: 33430354
- PMCID: PMC7825780
- DOI: 10.3390/cancers13020190
Cold-Shock Domains-Abundance, Structure, Properties, and Nucleic-Acid Binding
Abstract
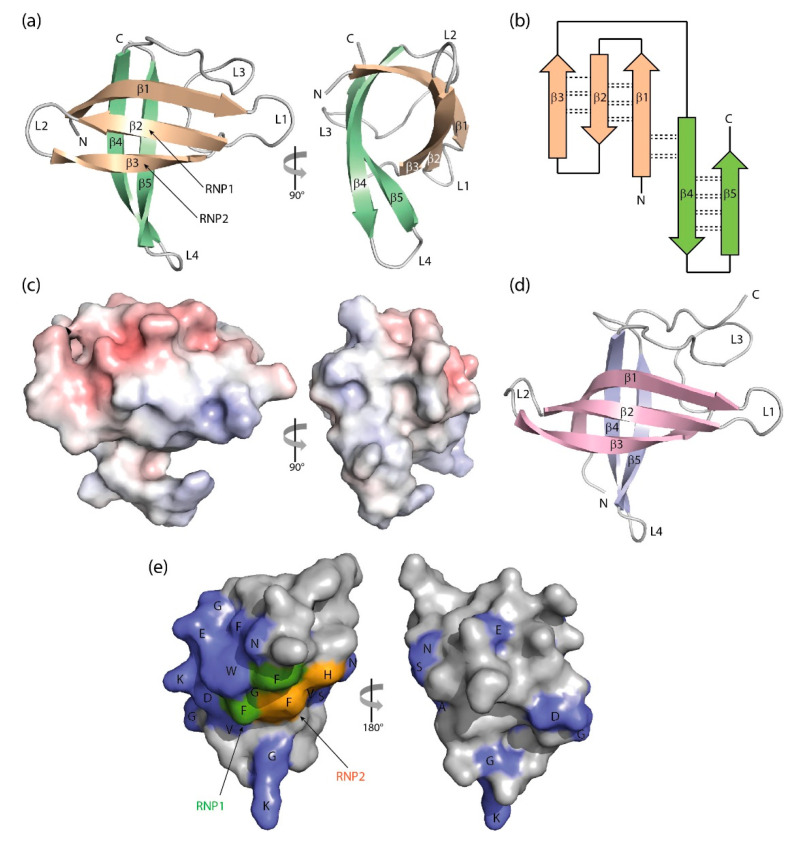
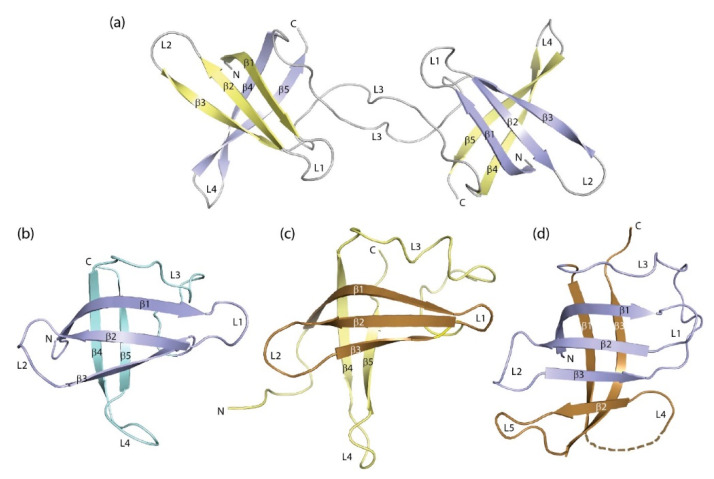
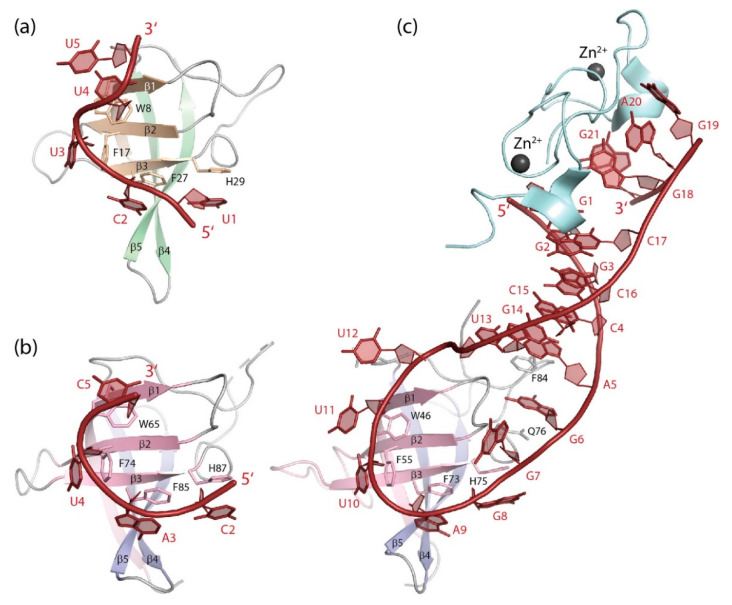
The cold-shock domain has a deceptively simple architecture but supports a complex biology. It is conserved from bacteria to man and has representatives in all kingdoms of life. Bacterial cold-shock proteins consist of a single cold-shock domain and some, but not all are induced by cold shock. Cold-shock domains in human proteins are often associated with natively unfolded protein segments and more rarely with other folded domains. Cold-shock proteins and domains share a five-stranded all-antiparallel β-barrel structure and a conserved surface that binds single-stranded nucleic acids, predominantly by stacking interactions between nucleobases and aromatic protein sidechains. This conserved binding mode explains the cold-shock domains' ability to associate with both DNA and RNA strands and their limited sequence selectivity. The promiscuous DNA and RNA binding provides a rationale for the ability of cold-shock domain-containing proteins to function in transcription regulation and DNA-damage repair as well as in regulating splicing, translation, mRNA stability and RNA sequestration.
Keywords: OB fold; RNA-binding domain; Y-box binding protein; cold-shock domain; cold-shock protein; domain fold; gene regulation; nucleic-acid binding; protein stability and folding; protein structure.
Conflict of interest statement
The authors declare no conflict of interest.
Figures



Similar articles
-
The solution structure and DNA-binding properties of the cold-shock domain of the human Y-box protein YB-1.J Mol Biol. 2002 Feb 15;316(2):317-26. doi: 10.1006/jmbi.2001.5334. J Mol Biol. 2002. PMID: 11851341
-
Universal nucleic acid-binding domain revealed by crystal structure of the B. subtilis major cold-shock protein.Nature. 1993 Jul 8;364(6433):164-8. doi: 10.1038/364164a0. Nature. 1993. PMID: 8321288
-
Common mode of DNA binding to cold shock domains. Crystal structure of hexathymidine bound to the domain-swapped form of a major cold shock protein from Bacillus caldolyticus.FEBS J. 2007 Mar;274(5):1265-79. doi: 10.1111/j.1742-4658.2007.05672.x. Epub 2007 Jan 31. FEBS J. 2007. PMID: 17266726
-
Comparison of structure, function and regulation of plant cold shock domain proteins to bacterial and animal cold shock domain proteins.BMB Rep. 2010 Jan;43(1):1-8. doi: 10.5483/bmbrep.2010.43.1.001. BMB Rep. 2010. PMID: 20132728 Review.
-
Y-box proteins combine versatile cold shock domains and arginine-rich motifs (ARMs) for pleiotropic functions in RNA biology.Biochem J. 2018 Sep 11;475(17):2769-2784. doi: 10.1042/BCJ20170956. Biochem J. 2018. PMID: 30206185 Review.
Cited by
-
DNA and RNA Binding Proteins: From Motifs to Roles in Cancer.Int J Mol Sci. 2022 Aug 18;23(16):9329. doi: 10.3390/ijms23169329. Int J Mol Sci. 2022. PMID: 36012592 Free PMC article. Review.
-
β-Barrels and Amyloids: Structural Transitions, Biological Functions, and Pathogenesis.Int J Mol Sci. 2021 Oct 20;22(21):11316. doi: 10.3390/ijms222111316. Int J Mol Sci. 2021. PMID: 34768745 Free PMC article. Review.
-
Transcriptome analyses reveal differences in the response to temperature in Florida and Northern largemouth bass (Micropterus spp.) during early life stages.PLoS One. 2025 Feb 18;20(2):e0317563. doi: 10.1371/journal.pone.0317563. eCollection 2025. PLoS One. 2025. PMID: 39964964 Free PMC article.
-
Carrot and stick: how RNase R contributes to function and destruction of the translation machinery.RNA Biol. 2025 Dec;22(1):1-22. doi: 10.1080/15476286.2025.2535846. Epub 2025 Jul 29. RNA Biol. 2025. PMID: 40734258 Free PMC article. Review.
-
Listeria monocytogenes Cold Shock Proteins: Small Proteins with A Huge Impact.Microorganisms. 2021 May 14;9(5):1061. doi: 10.3390/microorganisms9051061. Microorganisms. 2021. PMID: 34068949 Free PMC article. Review.
References
-
- Baltz A.G., Munschauer M., Schwanhausser B., Vasile A., Murakawa Y., Schueler M., Youngs N., Penfold-Brown D., Drew K., Milek M., et al. The mRNA-bound proteome and its global occupancy profile on protein-coding transcripts. Mol. Cell. 2012;46:674–690. doi: 10.1016/j.molcel.2012.05.021. - DOI - PubMed
Publication types
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
