

Regulation of a subset of release-ready vesicles by the presynaptic protein Mover
- PMID: 33431696
- PMCID: PMC7826340
- DOI: 10.1073/pnas.2022551118
Regulation of a subset of release-ready vesicles by the presynaptic protein Mover
Abstract
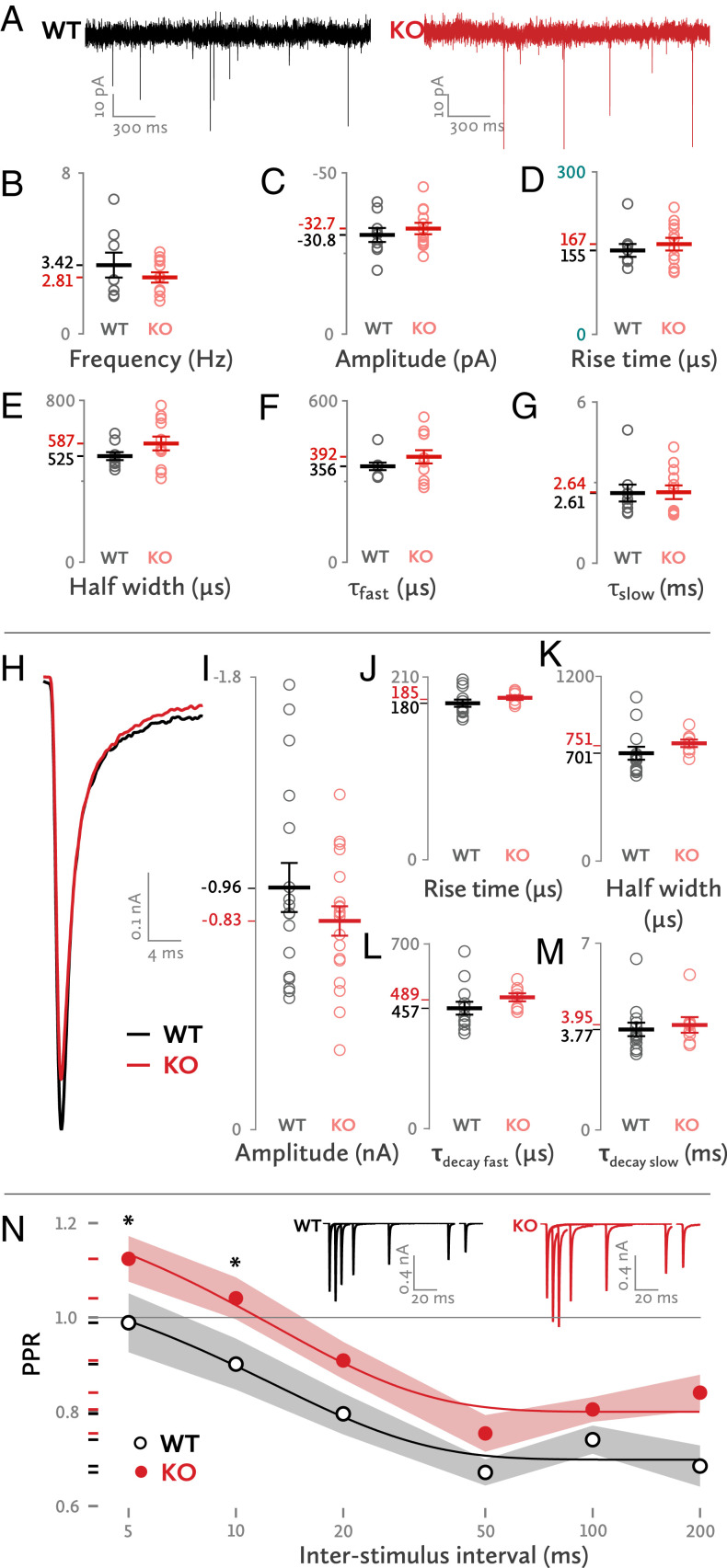
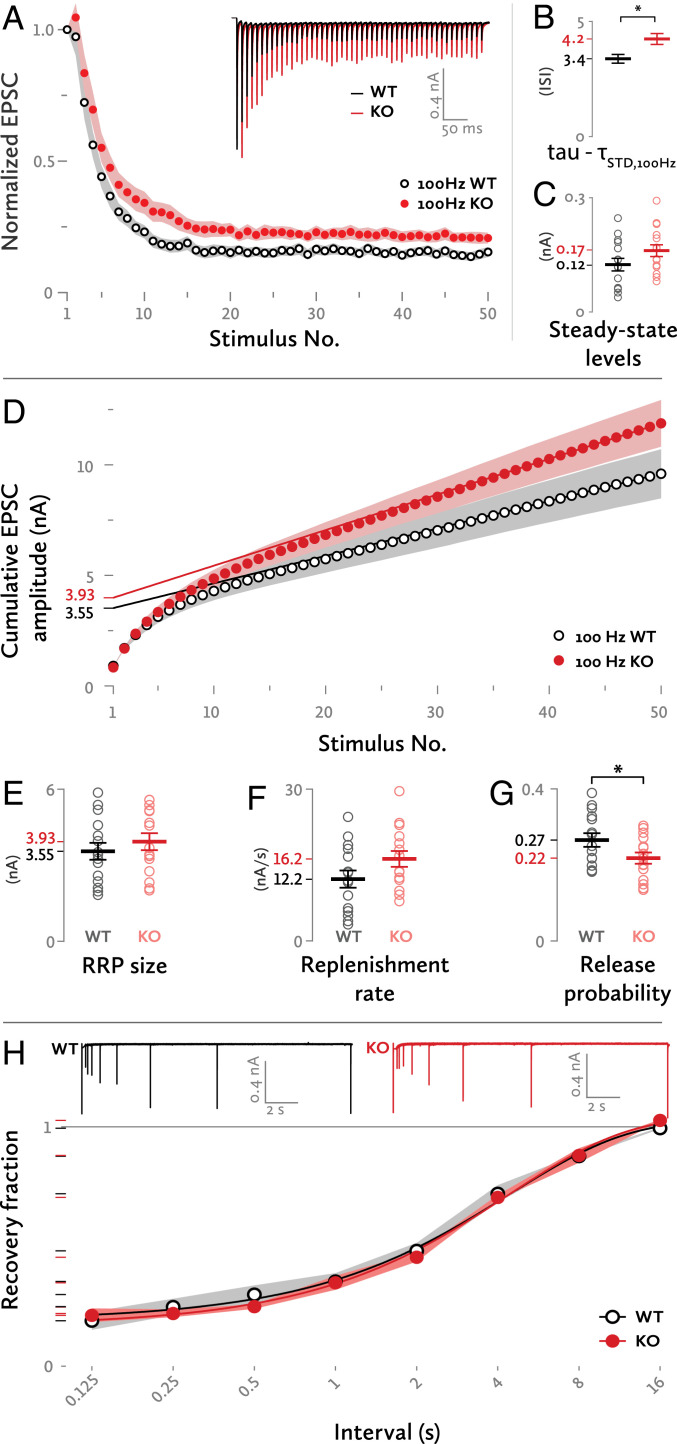
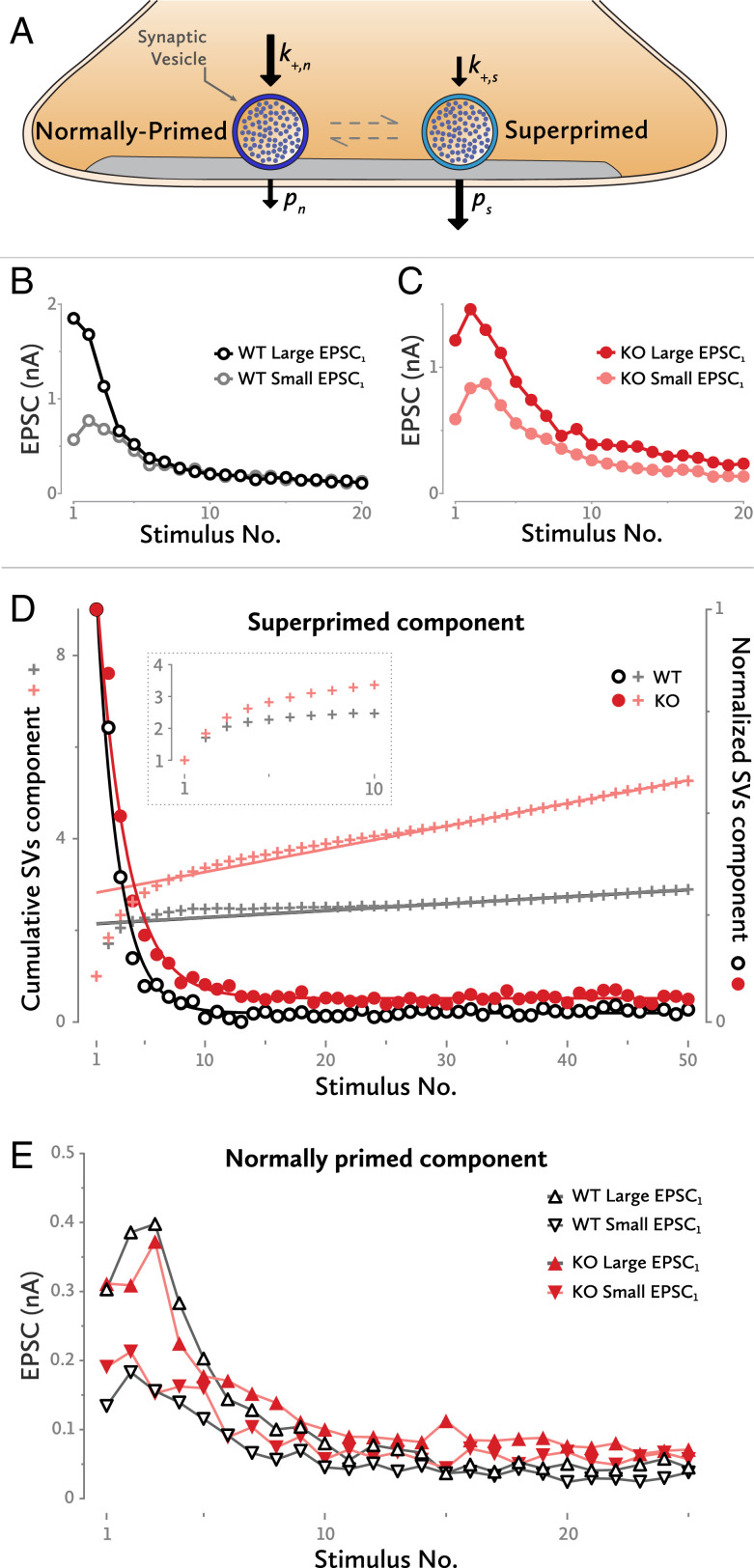
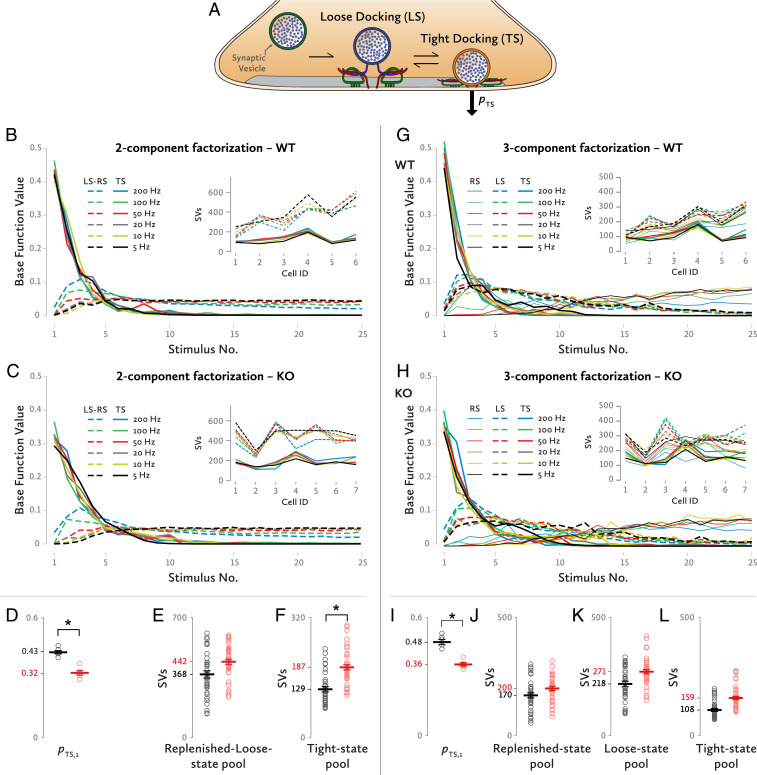
Neurotransmitter release occurs by regulated exocytosis from synaptic vesicles (SVs). Evolutionarily conserved proteins mediate the essential aspects of this process, including the membrane fusion step and priming steps that make SVs release-competent. Unlike the proteins constituting the core fusion machinery, the SV protein Mover does not occur in all species and all synapses. Its restricted expression suggests that Mover may modulate basic aspects of transmitter release and short-term plasticity. To test this hypothesis, we analyzed synaptic transmission electrophysiologically at the mouse calyx of Held synapse in slices obtained from wild-type mice and mice lacking Mover. Spontaneous transmission was unaffected, indicating that the basic release machinery works in the absence of Mover. Evoked release and vesicular release probability were slightly reduced, and the paired pulse ratio was increased in Mover knockout mice. To explore whether Mover's role is restricted to certain subpools of SVs, we analyzed our data in terms of two models of priming. A model assuming two SV pools in parallel showed a reduced release probability of so-called "superprimed vesicles" while "normally primed" ones were unaffected. For the second model, which holds that vesicles transit sequentially from a loosely docked state to a tightly docked state before exocytosis, we found that knocking out Mover selectively decreased the release probability of tight state vesicles. These results indicate that Mover regulates a subclass of primed SVs in the mouse calyx of Held.
Keywords: Mover; neurotransmission; presynaptic plasticity; synaptic vesicles; vesicle priming.
Copyright © 2021 the Author(s). Published by PNAS.
Conflict of interest statement
The authors declare no competing interest.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
