

Spatiotemporal variation in cell proliferation patterns during arthropod axial elongation
- PMID: 33431947
- PMCID: PMC7801698
- DOI: 10.1038/s41598-020-79373-0
Spatiotemporal variation in cell proliferation patterns during arthropod axial elongation
Abstract
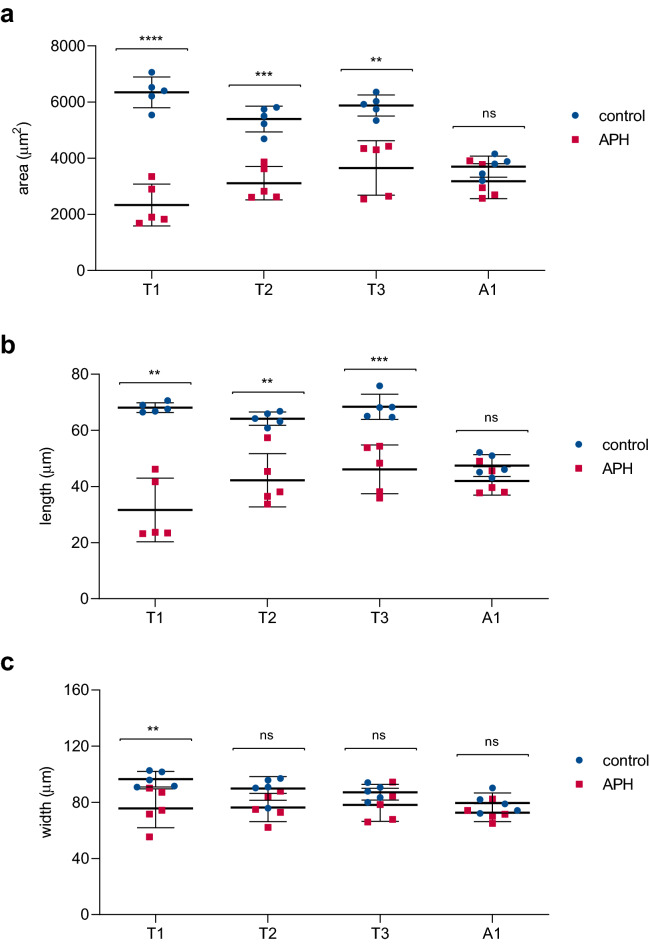
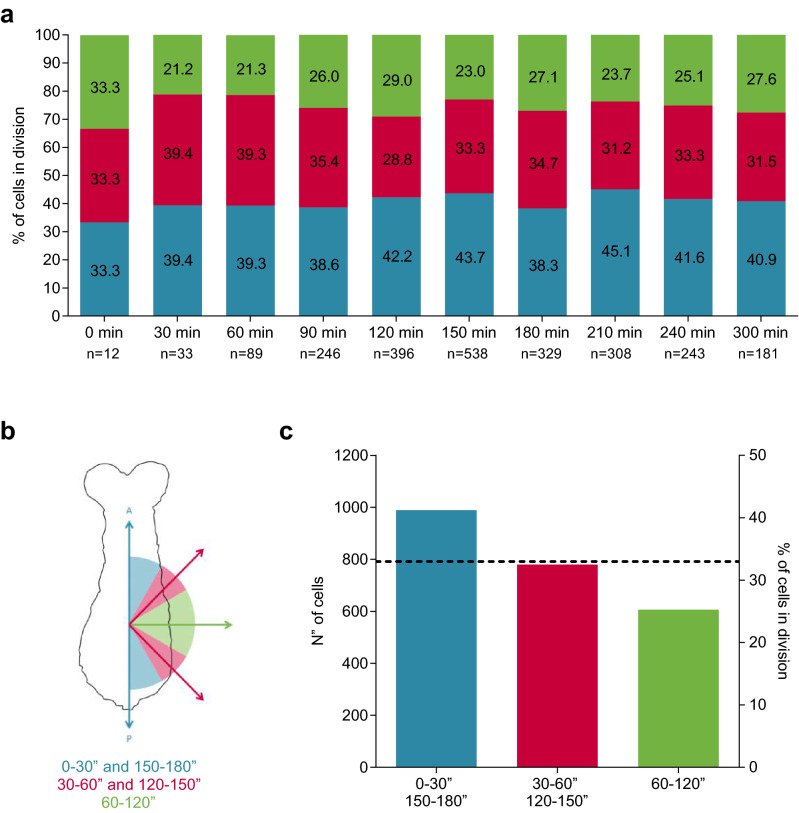
An elongated and segmented body plan is a common morphological characteristic of all arthropods and is probably responsible for their high adaptation ability to diverse environments. Most arthropods form their bodies by progressively adding segments, resembling vertebrate somitogenesis. This sequential segmentation relies on a molecular clock that operates in the posterior region of the elongating embryo that combines dynamically with cellular behaviors and tissue rearrangements, allowing the extension of the developing body along its main embryonic axis. Even though the molecular mechanisms involved in elongation and segment formation have been found to be conserved in a considerable degree, cellular processes such as cell division are quite variable between different arthropods. In this study, we show that cell proliferation in the beetle Tribolium castaneum has a nonuniform spatiotemporal patterning during axial elongation. We found that dividing cells are preferentially oriented along the anterior-posterior axis, more abundant and posteriorly localized during thoracic segments formation and that this cell proliferation peak was triggered at the onset of axis elongation. This raise in cell divisions, in turn, was correlated with an increase in the elongation rate, but not with changes in cell density. When DNA synthesis was inhibited over this period, both the area and length of thoracic segments were significantly reduced but not of the first abdominal segment. We discuss the variable participation that different cell division patterns and cell movements may have on arthropod posterior growth and their evolutionary contribution.
Conflict of interest statement
The authors declare no competing interests.
Figures



References
-
- Foe VE. Mitotic domains reveal early commitment of cells in Drosophila embryos. Development. 1989;107:1–22. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
