

Oxylipins are implicated as communication signals in tomato-root-knot nematode (Meloidogyne javanica) interaction
- PMID: 33431951
- PMCID: PMC7801703
- DOI: 10.1038/s41598-020-79432-6
Oxylipins are implicated as communication signals in tomato-root-knot nematode (Meloidogyne javanica) interaction
Abstract
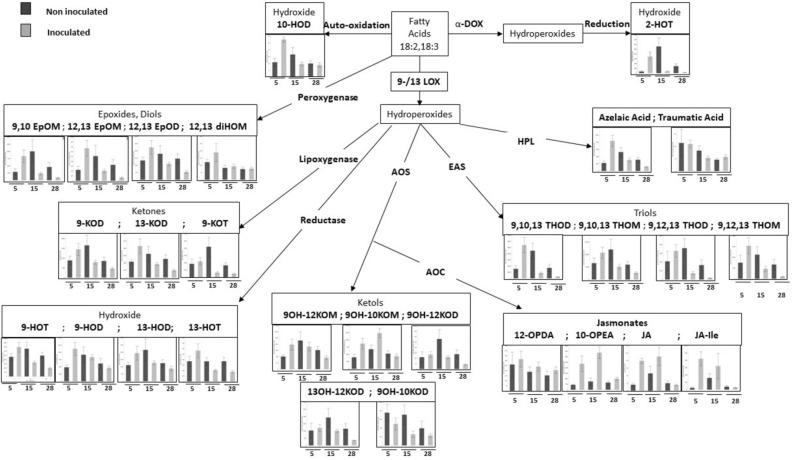
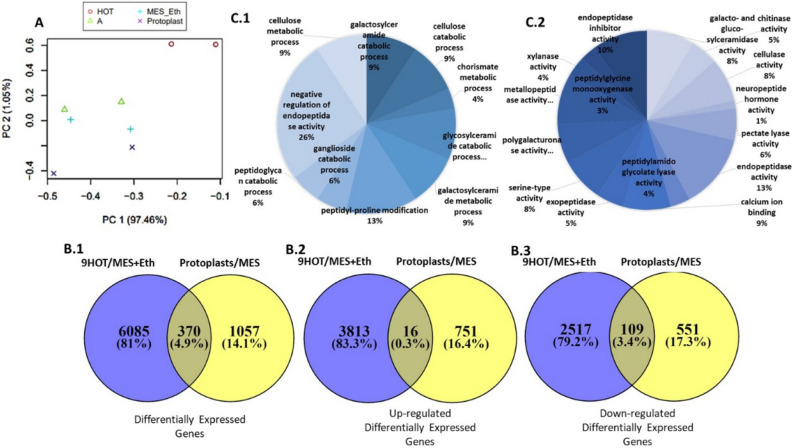
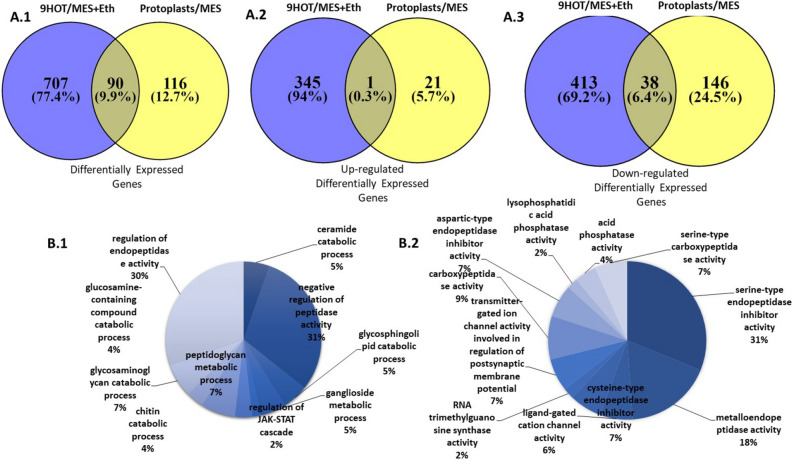
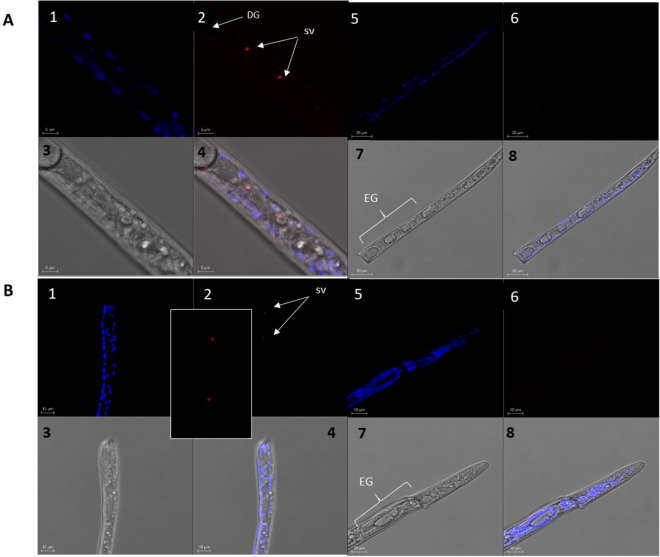
Throughout infection, plant-parasitic nematodes activate a complex host defense response that will regulate their development and aggressiveness. Oxylipins-lipophilic signaling molecules-are part of this complex, performing a fundamental role in regulating plant development and immunity. At the same time, the sedentary root-knot nematode Meloidogyne spp. secretes numerous effectors that play key roles during invasion and migration, supporting construction and maintenance of nematodes' feeding sites. Herein, comprehensive oxylipin profiling of tomato roots, performed using LC-MS/MS, indicated strong and early responses of many oxylipins following root-knot nematode infection. To identify genes that might respond to the lipidomic defense pathway mediated through oxylipins, RNA-Seq was performed by exposing Meloidogyne javanica second-stage juveniles to tomato protoplasts and the oxylipin 9-HOT, one of the early-induced oxylipins in tomato roots upon nematode infection. A total of 7512 differentially expressed genes were identified. To target putative effectors, we sought differentially expressed genes carrying a predicted secretion signal peptide. Among these, several were homologous with known effectors in other nematode species; other unknown, potentially secreted proteins may have a role as root-knot nematode effectors that are induced by plant lipid signals. These include effectors associated with distortion of the plant immune response or manipulating signal transduction mediated by lipid signals. Other effectors are implicated in cell wall degradation or ROS detoxification at the plant-nematode interface. Being an integral part of the plant's defense response, oxylipins might be placed as important signaling molecules underlying nematode parasitism.
Conflict of interest statement
The authors declare no competing interests.
Figures

References
-
- Nicol JM, et al. Genomics and Molecular Genetics of Plant–Nematode Interactions. Dordrecht: Springer; 2011. Current nematode threats to world agriculture; pp. 21–43.
-
- Singh S, Singh B, Singh AP. Nematodes: a threat to sustainability of agriculture. Procedia Environ. Sci. 2015;29:215–216. doi: 10.1016/j.proenv.2015.07.270. - DOI
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
