

R-2-hydroxyglutarate attenuates aerobic glycolysis in leukemia by targeting the FTO/m6A/PFKP/LDHB axis
- PMID: 33434505
- PMCID: PMC7935770
- DOI: 10.1016/j.molcel.2020.12.026
R-2-hydroxyglutarate attenuates aerobic glycolysis in leukemia by targeting the FTO/m6A/PFKP/LDHB axis
Abstract
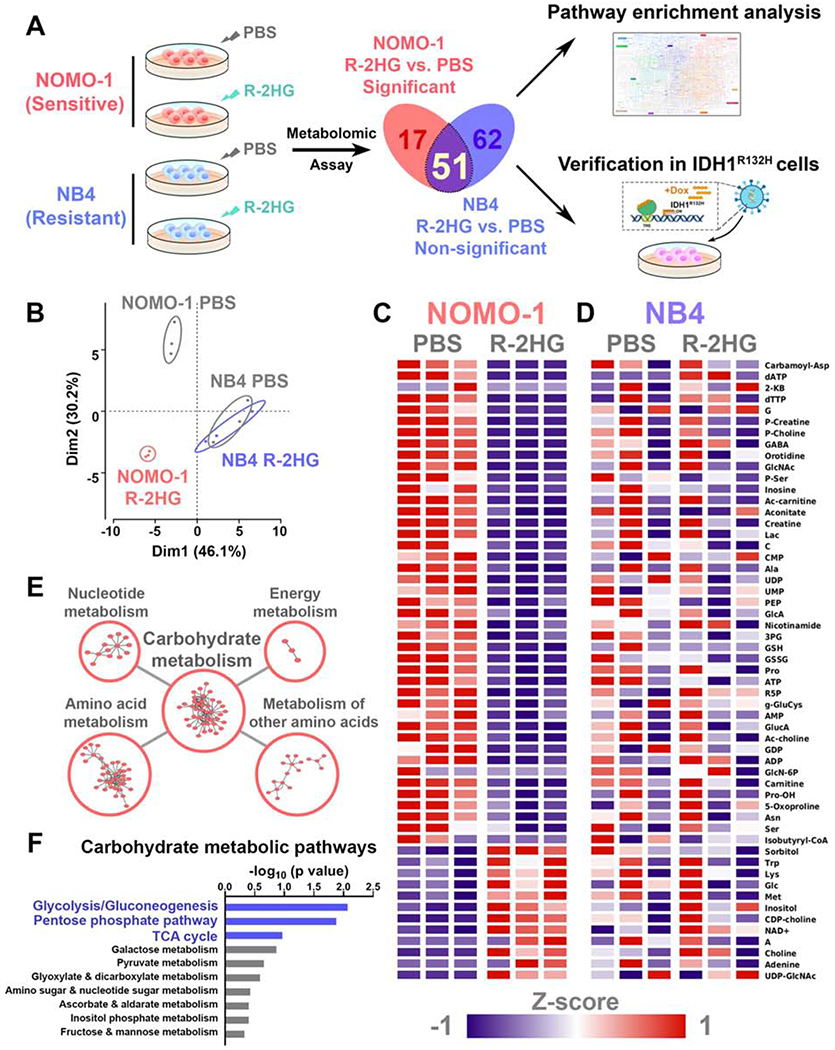
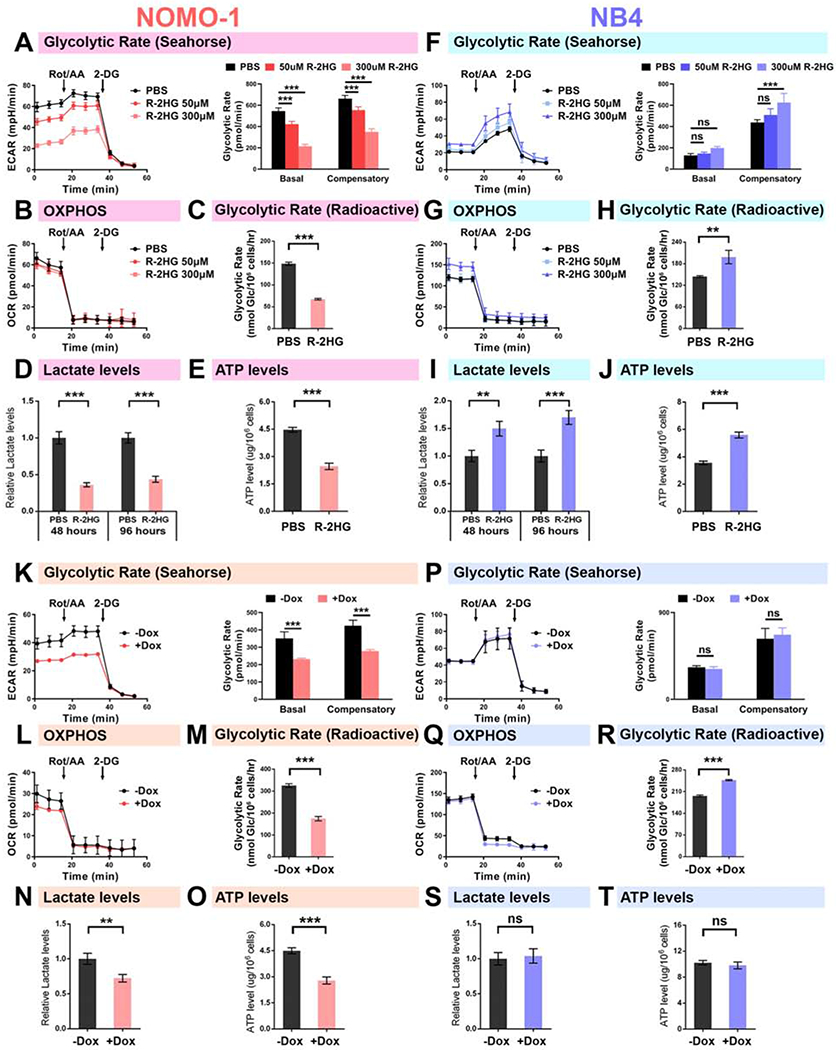
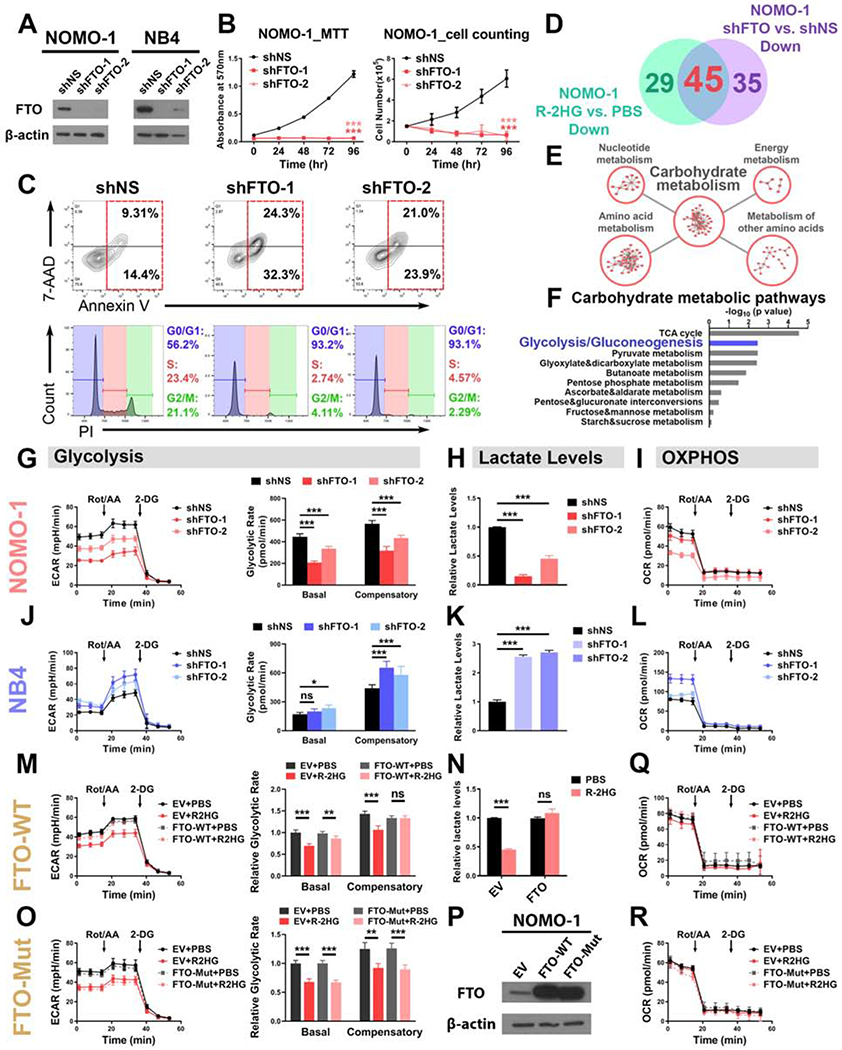
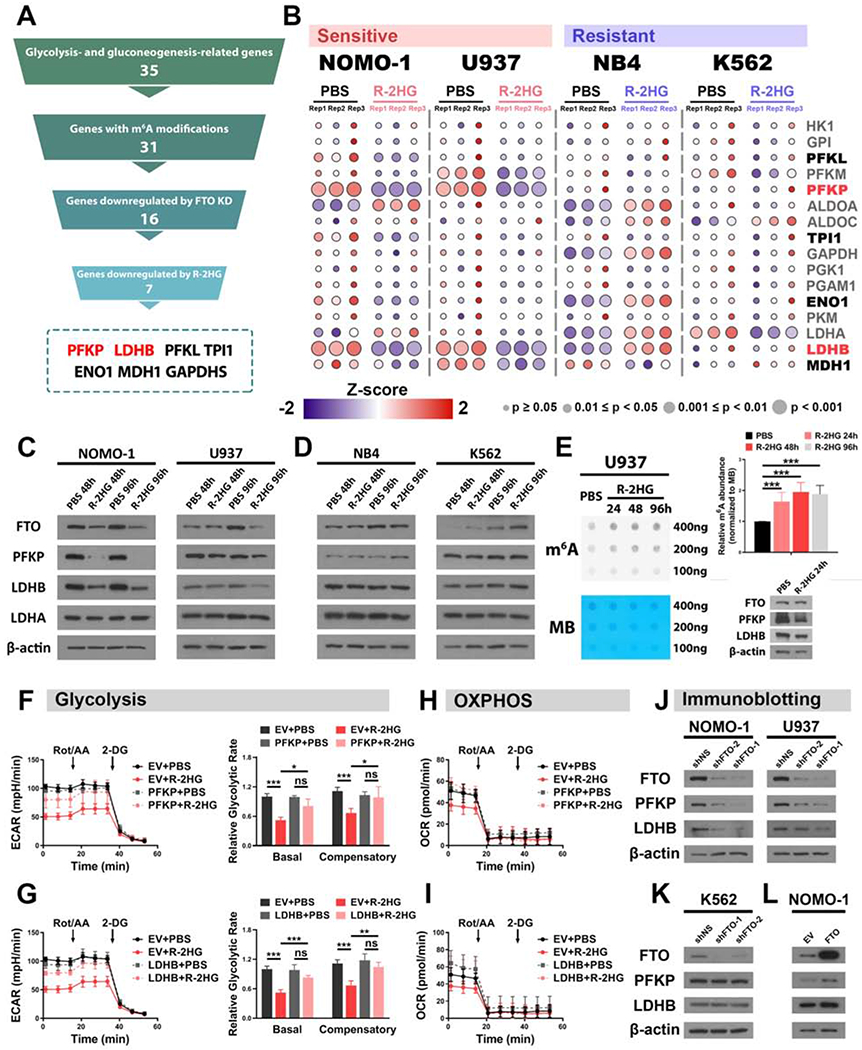
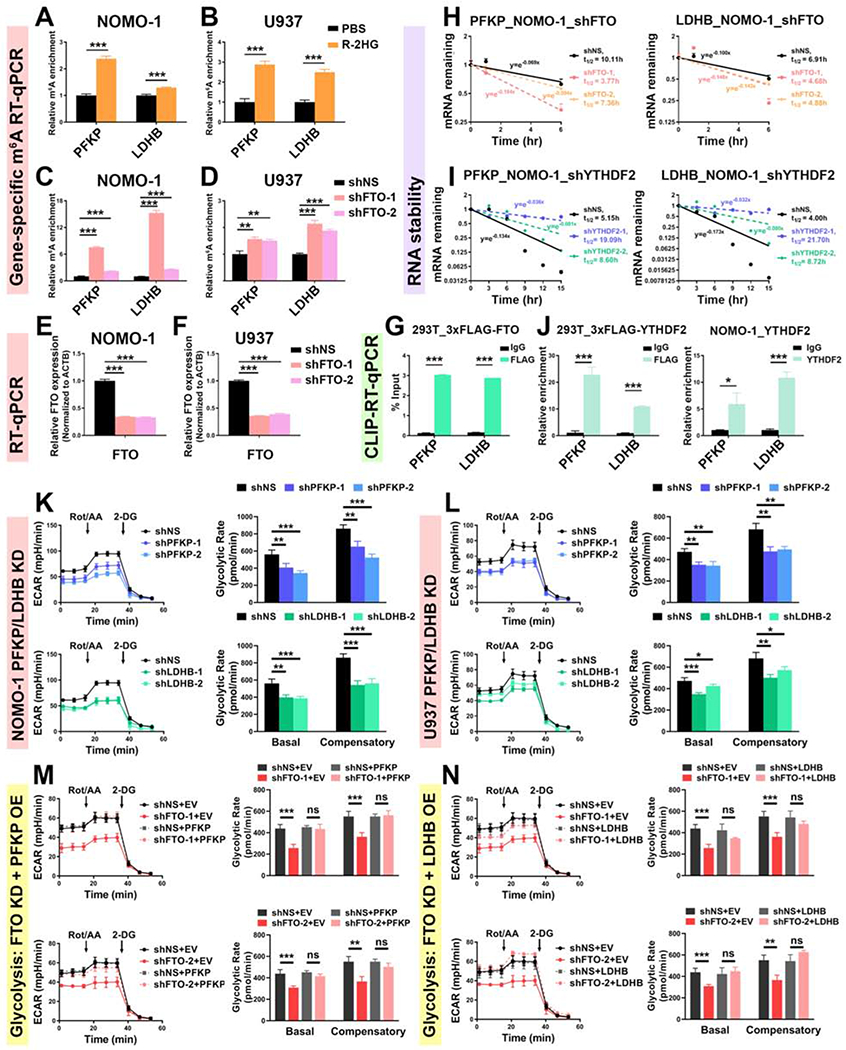
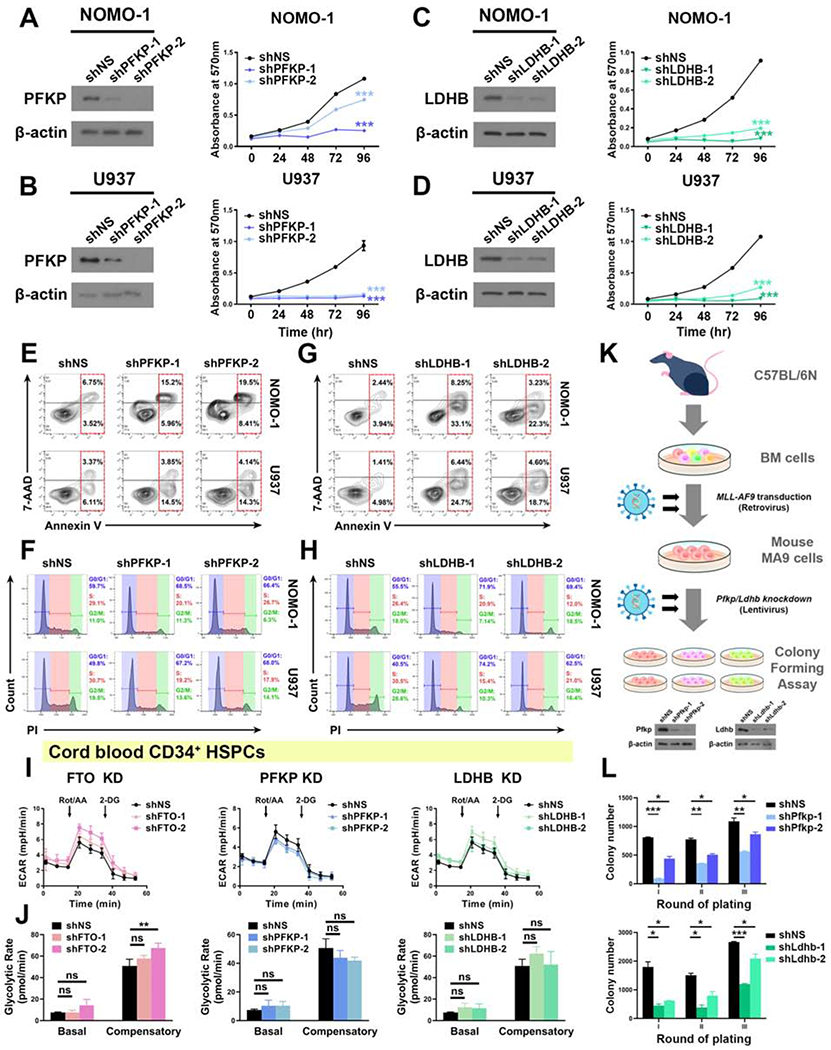
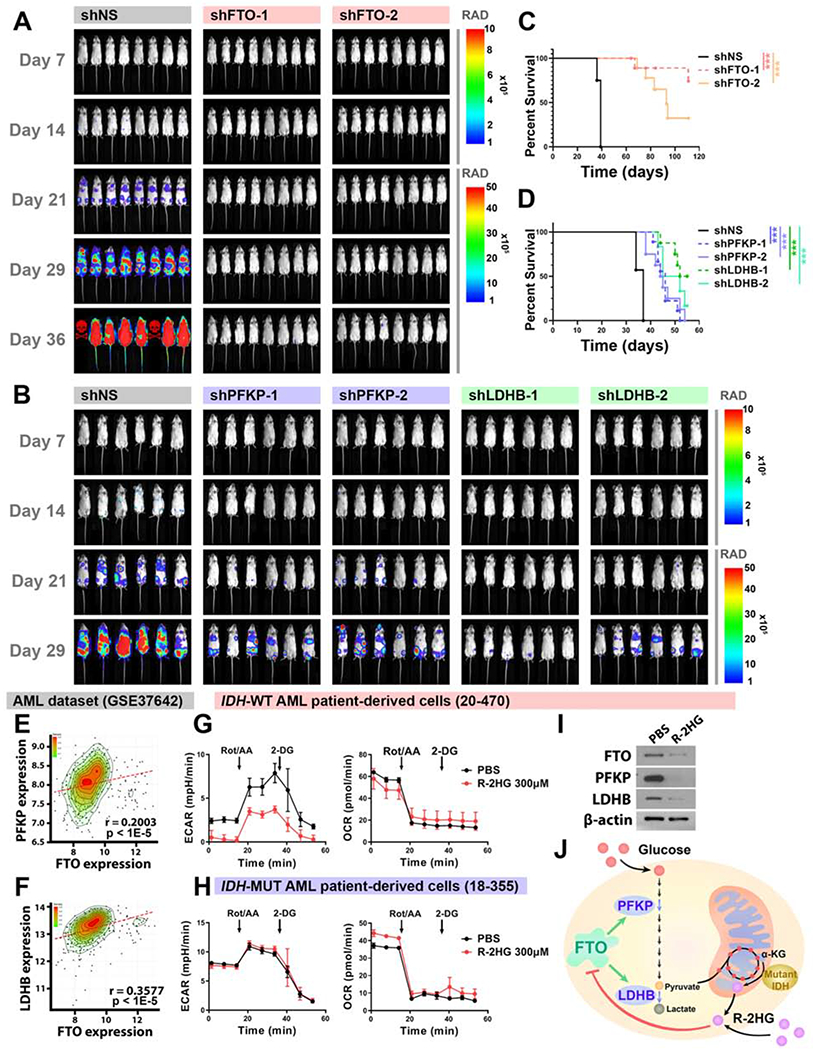
R-2-hydroxyglutarate (R-2HG), a metabolite produced by mutant isocitrate dehydrogenases (IDHs), was recently reported to exhibit anti-tumor activity. However, its effect on cancer metabolism remains largely elusive. Here we show that R-2HG effectively attenuates aerobic glycolysis, a hallmark of cancer metabolism, in (R-2HG-sensitive) leukemia cells. Mechanistically, R-2HG abrogates fat-mass- and obesity-associated protein (FTO)/N6-methyladenosine (m6A)/YTH N6-methyladenosine RNA binding protein 2 (YTHDF2)-mediated post-transcriptional upregulation of phosphofructokinase platelet (PFKP) and lactate dehydrogenase B (LDHB) (two critical glycolytic genes) expression and thereby suppresses aerobic glycolysis. Knockdown of FTO, PFKP, or LDHB recapitulates R-2HG-induced glycolytic inhibition in (R-2HG-sensitive) leukemia cells, but not in normal CD34+ hematopoietic stem/progenitor cells, and inhibits leukemogenesis in vivo; conversely, their overexpression reverses R-2HG-induced effects. R-2HG also suppresses glycolysis and downregulates FTO/PFKP/LDHB expression in human primary IDH-wild-type acute myeloid leukemia (AML) cells, demonstrating the clinical relevance. Collectively, our study reveals previously unrecognized effects of R-2HG and RNA modification on aerobic glycolysis in leukemia, highlighting the therapeutic potential of targeting cancer epitranscriptomics and metabolism.
Keywords: FTO; LDHB; N(6)-methyladenosine (m(6)A) modification; PFKP; R-2HG; RNA stability; YTHDF2; cancer metabolism; glycolysis; leukemia.
Copyright © 2020 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests J.C. is a scientific founder of Genovel Biotech Corp. and holds equities with the company.
Figures
References
-
- Bonnet S, Archer SL, Allalunis-Turner J, Haromy A, Beaulieu C, Thompson R, Lee CT, Lopaschuk GD, Puttagunta L, Bonnet S, et al. (2007). A mitochondria-K+ channel axis is suppressed in cancer and its normalization promotes apoptosis and inhibits cancer growth. Cancer Cell 11, 37–51. - PubMed
-
- Brisson L, Banski P, Sboarina M, Dethier C, Danhier P, Fontenille MJ, Van Hee VF, Vazeille T, Tardy M, Falces J, et al. (2016). Lactate Dehydrogenase B Controls Lysosome Activity and Autophagy in Cancer. Cancer Cell 30, 418–431. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- R01 CA236399/CA/NCI NIH HHS/United States
- R01 DK124116/DK/NIDDK NIH HHS/United States
- R01 CA214965/CA/NCI NIH HHS/United States
- R01 CA172558/CA/NCI NIH HHS/United States
- R01 CA169458/CA/NCI NIH HHS/United States
- R35 CA197628/CA/NCI NIH HHS/United States
- R01 CA243386/CA/NCI NIH HHS/United States
- HHMI/Howard Hughes Medical Institute/United States
- T32 CA186895/CA/NCI NIH HHS/United States
- R01 CA213138/CA/NCI NIH HHS/United States
- R01 CA157644/CA/NCI NIH HHS/United States
- R01 CA211614/CA/NCI NIH HHS/United States
- U10 CA180827/CA/NCI NIH HHS/United States
- R01 CA137060/CA/NCI NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
