

Recurrent network dynamics shape direction selectivity in primary auditory cortex
- PMID: 33436635
- PMCID: PMC7804939
- DOI: 10.1038/s41467-020-20590-6
Recurrent network dynamics shape direction selectivity in primary auditory cortex
Abstract
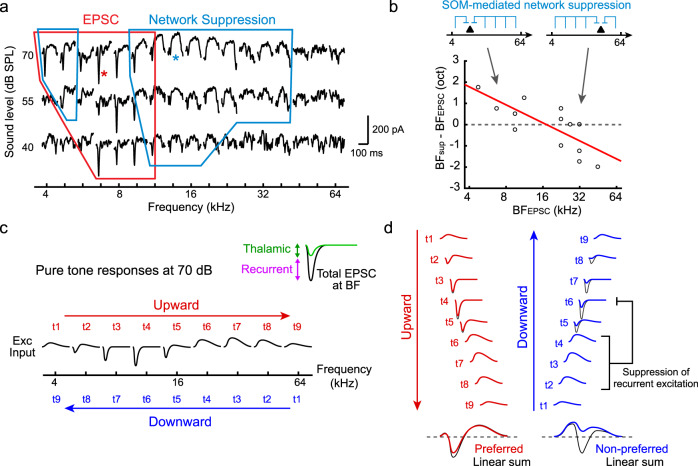
Detecting the direction of frequency modulation (FM) is essential for vocal communication in both animals and humans. Direction-selective firing of neurons in the primary auditory cortex (A1) has been classically attributed to temporal offsets between feedforward excitatory and inhibitory inputs. However, it remains unclear how cortical recurrent circuitry contributes to this computation. Here, we used two-photon calcium imaging and whole-cell recordings in awake mice to demonstrate that direction selectivity is not caused by temporal offsets between synaptic currents, but by an asymmetry in total synaptic charge between preferred and non-preferred directions. Inactivation of cortical somatostatin-expressing interneurons (SOM cells) reduced direction selectivity, revealing its cortical contribution. Our theoretical models showed that charge asymmetry arises due to broad spatial topography of SOM cell-mediated inhibition which regulates signal amplification in strongly recurrent circuitry. Together, our findings reveal a major contribution of recurrent network dynamics in shaping cortical tuning to behaviorally relevant complex sounds.
Conflict of interest statement
The authors declare no competing interests.
Figures






References
-
- Griffin, D. R. Listening in the Dark, The Acoustic Orientation of Bats and Men. (Yale University Press, 1958).
-
- Reichardt, W. Sensory Communication. (MIT Press, 1961).
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
