

Enzastaurin enhances ATRA-induced differentiation of acute myeloid leukemia cells
- PMID: 33437364
- PMCID: PMC7791522
Enzastaurin enhances ATRA-induced differentiation of acute myeloid leukemia cells
Abstract
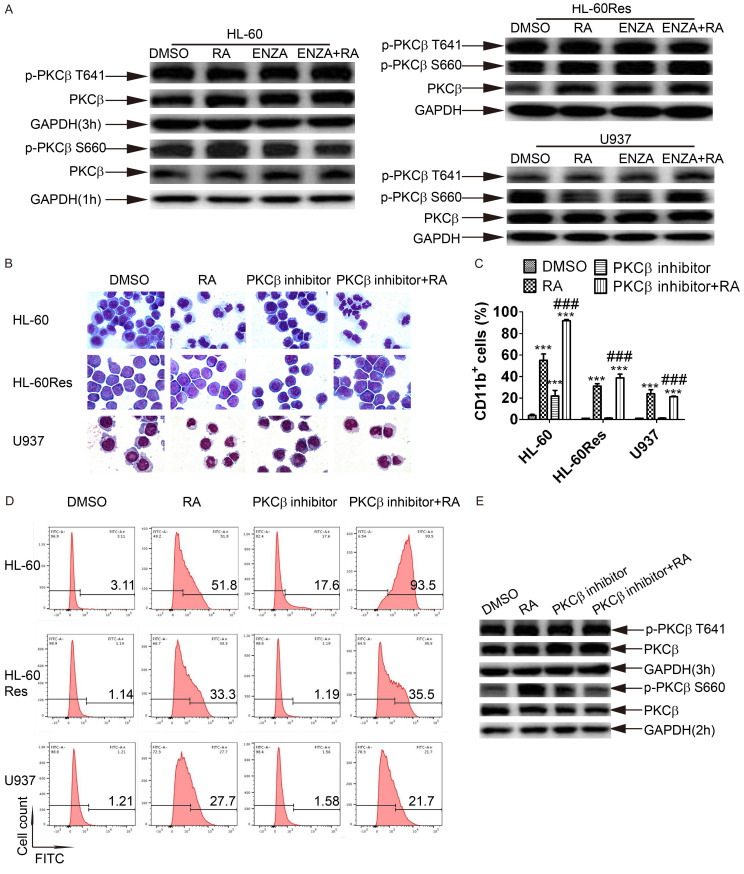
All-trans retinoic acid (ATRA) is considered to be the sole clinically-useful differentiating agent in the treatment of acute myeloid leukemia (AML). However, ATRA has been effective only in acute promyelocytic leukemia (APL) but not other subtypes of AML. Therefore, discovering strategies to sensitize cells to ATRA may lead to the development of ATRA-based treatments in non-APL AML patients. In the present study, a clinically-achievable concentration of enzastaurin enhanced ATRA-induced differentiation in AML cell lines, HL-60 and U937 as well as non-APL AML primary cells. Furthermore, it also restored ATRA sensitivity in ATRA-resistant cell line, HL-60Res. Mechanistically, in all these cell lines, enzastaurin-ATRA (enz-ATRA) co-treatment enhanced the protein levels of PU.1, CCAAT/enhancer-binding protein β (C/EBPβ) and C/EBPε. The activity of protein kinase C β (PKCβ) was suppressed by enz-ATRA treatment in HL-60 and HL-60Res cells. However, another PKCβ-selective inhibitor mimicked the cellular and molecular effects of enzastaurin only in HL-60 cells. Furthermore, in U937 cells, enz-ATRA activated MEK and ERK, and a MEK-specific inhibitor suppressed enz-ATRA-triggered differentiation and reduced the protein levels of PU.1, C/EBPβ and C/EBPε. Enz-ATRA activated Akt in HL-60 and HL-60Res cells. However, an Akt inhibitor blocked enz-ATRA-triggered differentiation and restored the protein levels of PU.1, C/EBPβ and C/EBPε only in HL-60Res cells. Therefore, PKCβ inhibition, MEK/ERK and Akt activation were involved in enz-ATRA-induced differentiation in HL-60, U937 and HL-60Res cells, respectively, via modulation of the protein levels of C/EBPβ, C/EBPε and PU.1. Taken together, our findings may help to guide novel therapeutic strategies for AML patients.
Keywords: Acute myeloid leukemia; all-trans retinoic acid; differentiation; enzastaurin.
AJTR Copyright © 2020.
Conflict of interest statement
None.
Figures






Similar articles
-
Combination of midostaurin and ATRA exerts dose-dependent dual effects on acute myeloid leukemia cells with wild type FLT3.BMC Cancer. 2022 Jul 9;22(1):749. doi: 10.1186/s12885-022-09828-2. BMC Cancer. 2022. PMID: 35810308 Free PMC article.
-
Combination of enzastaurin and ATRA exerts dose-dependent dual effects on ATRA-resistant acute promyelocytic leukemia cells.Am J Cancer Res. 2019 May 1;9(5):906-926. eCollection 2019. Am J Cancer Res. 2019. PMID: 31218101 Free PMC article.
-
Trametinib enhances ATRA-induced differentiation in AML cells.Leuk Lymphoma. 2021 Dec;62(14):3361-3372. doi: 10.1080/10428194.2021.1961231. Epub 2021 Aug 6. Leuk Lymphoma. 2021. PMID: 34355652
-
Reprogramming acute myeloid leukemia into sensitivity for retinoic-acid-driven differentiation.Exp Hematol. 2017 Aug;52:12-23. doi: 10.1016/j.exphem.2017.04.007. Epub 2017 Apr 27. Exp Hematol. 2017. PMID: 28456748 Review.
-
All-trans retinoic acid and hematopoietic growth factors regulating the growth and differentiation of blast progenitors in acute promyelocytic leukemia.Leuk Lymphoma. 1999 Apr;33(3-4):267-80. doi: 10.3109/10428199909058426. Leuk Lymphoma. 1999. PMID: 10221506 Review.
Cited by
-
FTO effects the proliferation, invasion, and glycolytic metabolism of colon cancer by regulating PKM2.J Cancer Res Clin Oncol. 2025 Jan 16;151(1):36. doi: 10.1007/s00432-024-06073-x. J Cancer Res Clin Oncol. 2025. PMID: 39820532 Free PMC article.
-
Combination of Ethacrynic Acid and ATRA Triggers Differentiation and/or Apoptosis of Acute Myeloid Leukemia Cells through ROS.Anticancer Agents Med Chem. 2024;24(6):412-422. doi: 10.2174/0118715206273000231211092743. Anticancer Agents Med Chem. 2024. PMID: 38204257
-
Unlocking therapeutic potential: integration of drug repurposing and immunotherapy for various disease targeting.Am J Transl Res. 2023 Aug 15;15(8):4984-5006. eCollection 2023. Am J Transl Res. 2023. PMID: 37692967 Free PMC article. Review.
-
Combination of midostaurin and ATRA exerts dose-dependent dual effects on acute myeloid leukemia cells with wild type FLT3.BMC Cancer. 2022 Jul 9;22(1):749. doi: 10.1186/s12885-022-09828-2. BMC Cancer. 2022. PMID: 35810308 Free PMC article.
References
-
- Döhner H, Weisdorf DJ, Bloomfield CD. Acute myeloid leukemia. N Engl J Med. 2015;373:1136–1152. - PubMed
-
- Huang ME, Ye YC, Chen SR, Chai JR, Lu JX, Zhoa L, Gu LJ, Wang ZY. Use of all-trans retinoic acid in the treatment of acute promyelocytic leukemia. Blood. 1988;72:567–572. - PubMed
-
- Tallman MS. Differentiating therapy with all-trans retinoic acid in acute myeloid leukemia. Leukemia. 1996;10(Suppl 1):S12–S15. - PubMed
-
- Schlenk RF, Döhner K, Kneba M, Götze K, Hartmann F, Del Valle F, Kirchen H, Koller E, Fischer JT, Bullinger L, Habdank M, Späth D, Groner S, Krebs B, Kayser S, Corbacioglu A, Anhalt A, Benner A, Fröhling S, Döhner H. Gene mutations and response to treatment with all-trans retinoic acid in elderly patients with acute myeloid leukemia. Results from the AMLSG Trial AML HD98B. Haematologica. 2009;94:54–60. - PMC - PubMed
LinkOut - more resources
Full Text Sources
Miscellaneous