

Chemosensory Neurons Modulate the Response to Oomycete Recognition in Caenorhabditis elegans
- PMID: 33440164
- PMCID: PMC7809619
- DOI: 10.1016/j.celrep.2020.108604
Chemosensory Neurons Modulate the Response to Oomycete Recognition in Caenorhabditis elegans
Abstract
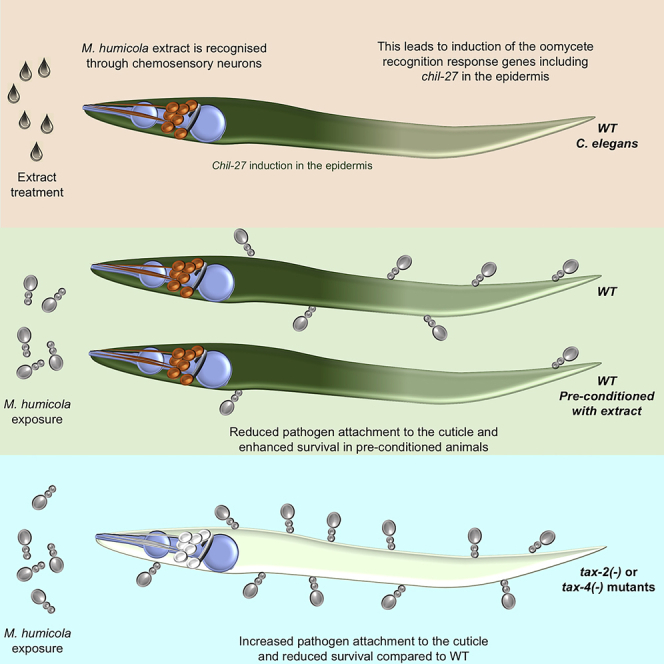
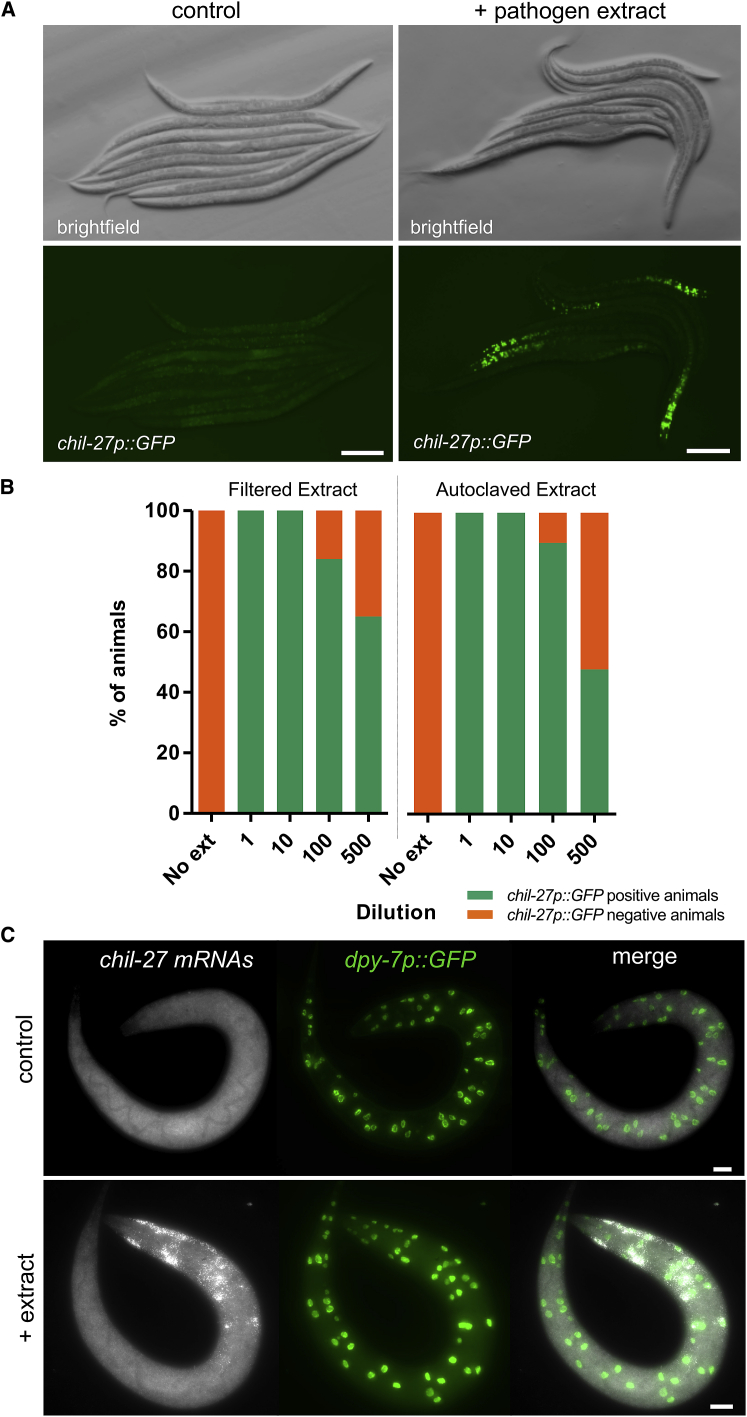
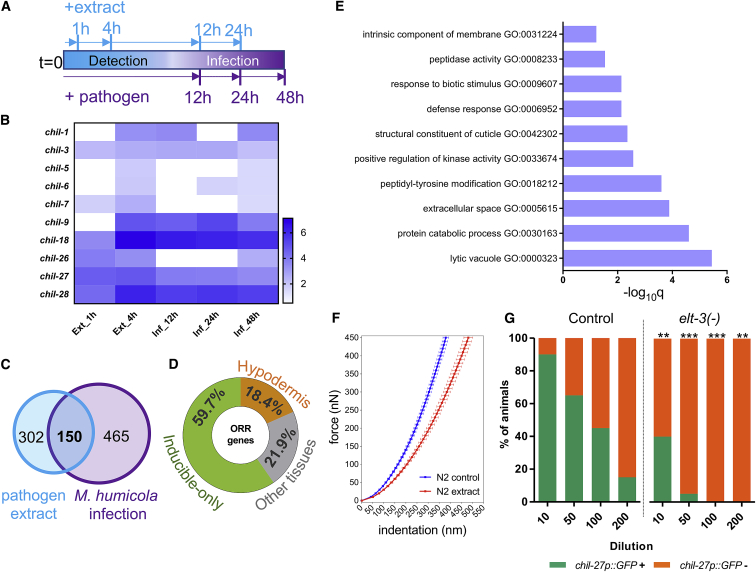
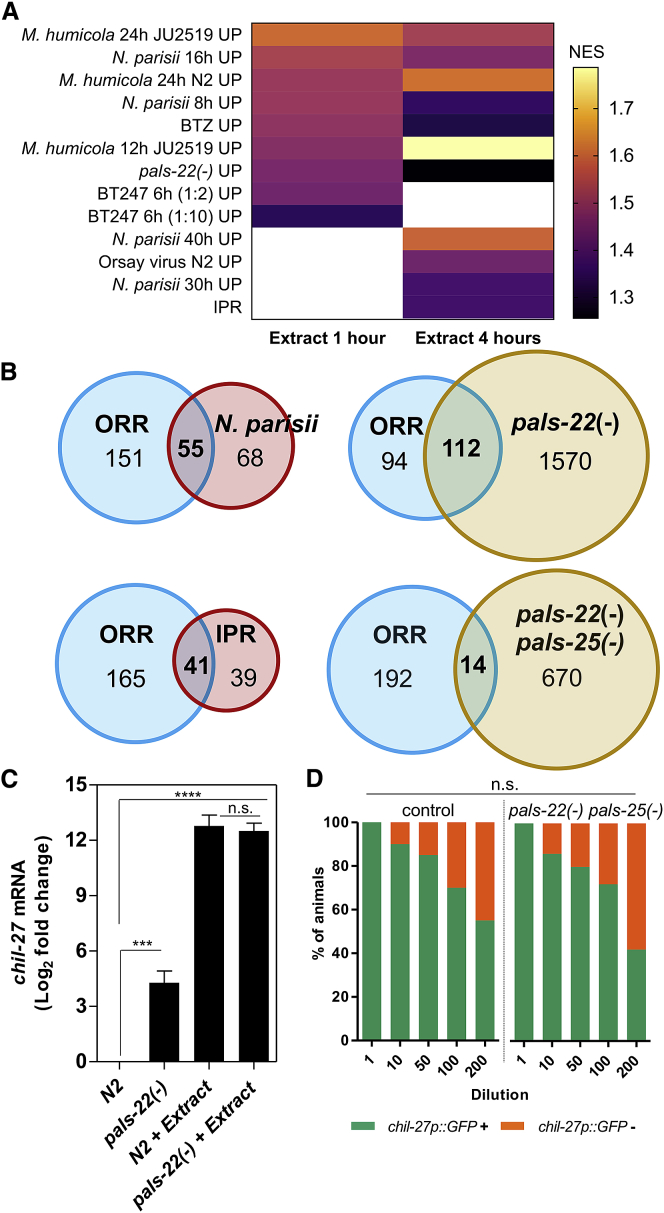
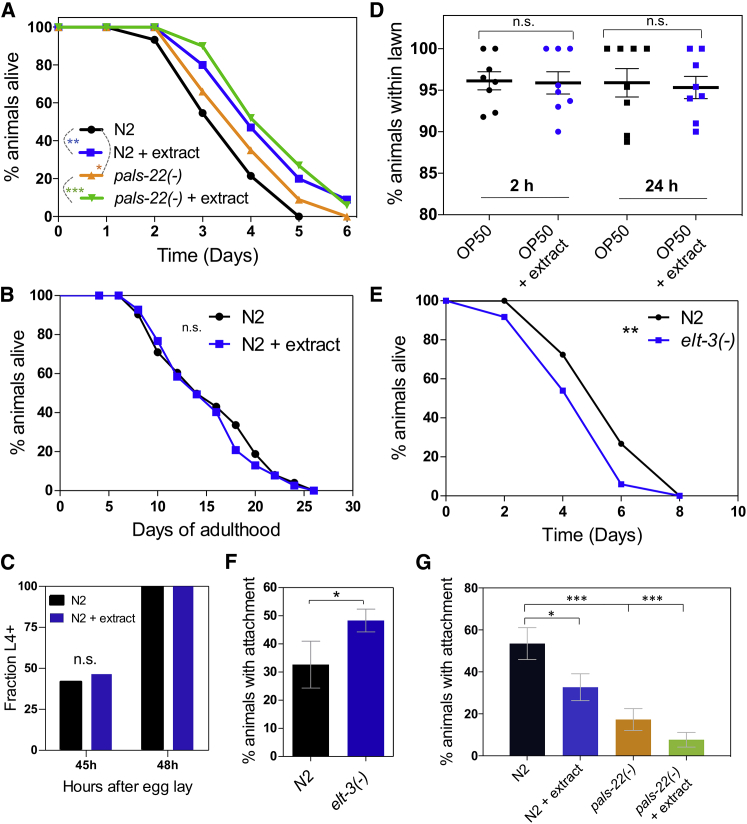
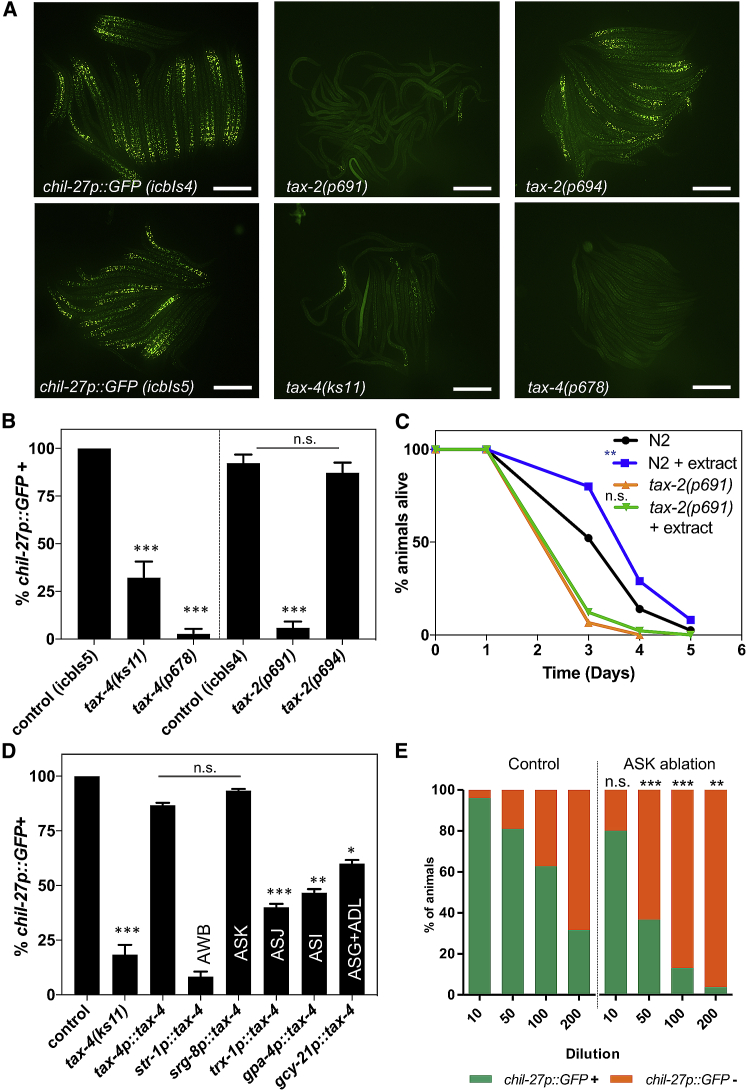
Understanding how animals detect and respond to pathogen threats is central to dissecting mechanisms of host immunity. The oomycetes represent a diverse eukaryotic group infecting various hosts from nematodes to humans. We have previously shown that Caenorhabditis elegans mounts a defense response consisting of the induction of chitinase-like (chil) genes in the epidermis to combat infection by its natural oomycete pathogen Myzocytiopsis humicola. We provide here evidence that C. elegans can sense the oomycete by detecting an innocuous extract derived from animals infected with M. humicola. The oomycete recognition response (ORR) leads to changes in the cuticle and reduction in pathogen attachment, thereby increasing animal survival. We also show that TAX-2/TAX-4 function in chemosensory neurons is required for the induction of chil-27 in the epidermis in response to extract exposure. Our findings highlight that neuron-to-epidermis communication may shape responses to oomycete recognition in animal hosts.
Keywords: Caenorhabditis elegans; Myzocytiopsis humicola; chitinase-like genes; cross-tissue signaling; cuticle; hypodermis; innate immunity; oomycete; pals; pathogen recognition.
Copyright © 2020 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of Interests The authors declare no competing interests.
Figures
References
-
- Akira S., Uematsu S., Takeuchi O. Pathogen recognition and innate immunity. Cell. 2006;124:783–801. - PubMed
-
- Beakes G.W., Glockling S.L., Sekimoto S. The evolutionary phylogeny of the oomycete “fungi”. Protoplasma. 2012;249:3–19. - PubMed
-
- Bigg H.F., Wait R., Rowan A.D., Cawston T.E. The mammalian chitinase-like lectin, YKL-40, binds specifically to type I collagen and modulates the rate of type I collagen fibril formation. J. Biol. Chem. 2006;281:21082–21095. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
