

The Involvement of Innate and Adaptive Immunity in the Initiation and Perpetuation of Sjögren's Syndrome
- PMID: 33440862
- PMCID: PMC7826728
- DOI: 10.3390/ijms22020658
The Involvement of Innate and Adaptive Immunity in the Initiation and Perpetuation of Sjögren's Syndrome
Abstract
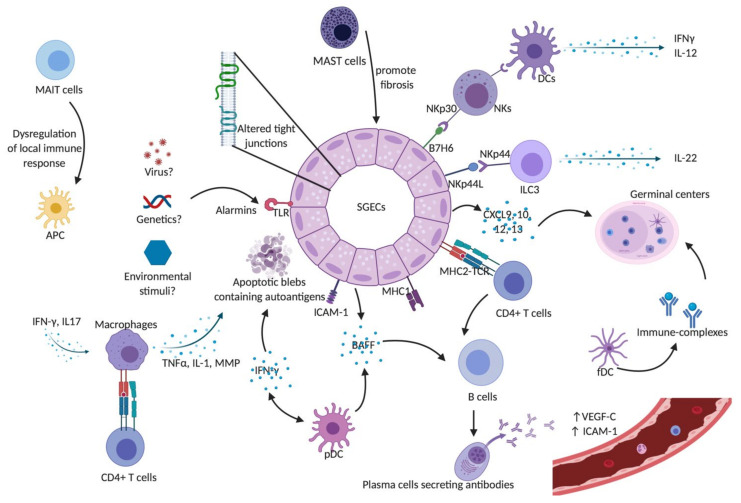
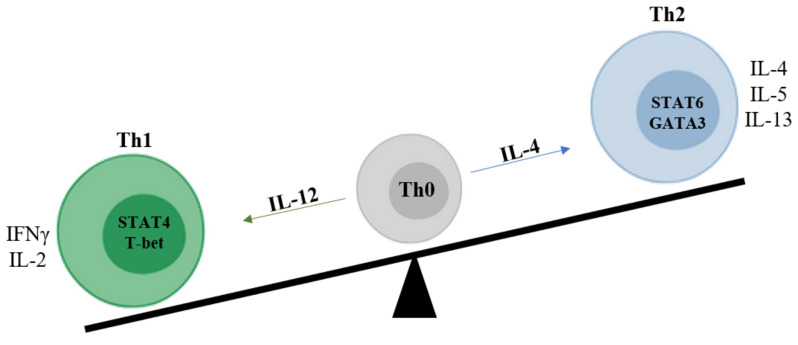
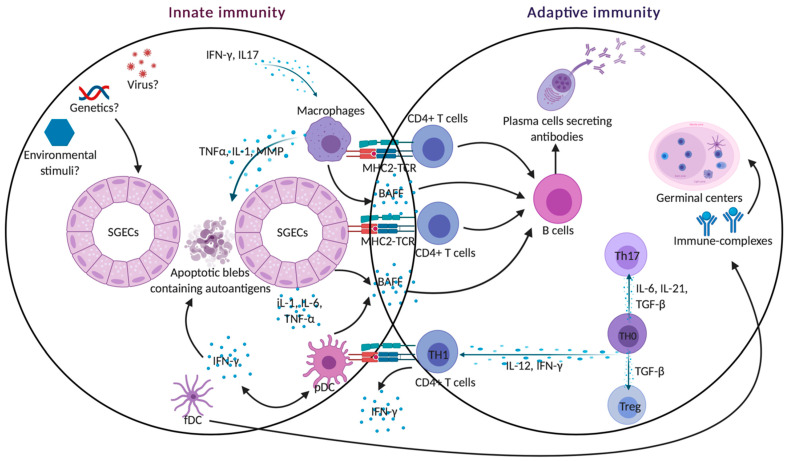
Sjogren's syndrome (SS) is a chronic autoimmune disease characterized by the infiltration of exocrine glands including salivary and lachrymal glands responsible for the classical dry eyes and mouth symptoms (sicca syndrome). The spectrum of disease manifestations stretches beyond the classical sicca syndrome with systemic manifestations including arthritis, interstitial lung involvement, and neurological involvement. The pathophysiology underlying SS is not well deciphered, but several converging lines of evidence have supported the conjuncture of different factors interplaying together to foster the initiation and perpetuation of the disease. The innate and adaptive immune system play a cardinal role in this process. In this review, we discuss the inherent parts played by both the innate and adaptive immune system in the pathogenesis of SS.
Keywords: b cells; epithelial cells; innate immunity; lymphocytes; sjogren’s syndrome; t cells.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Fox R.I., Kang H.I. Pathogenesis of Sjogren’s syndrome. Rheum. Dis. Clin. N. Am. 1992;18:517–538. - PubMed
-
- Fox R.I., Michelson P. Approaches to the treatment of Sjogren’s syndrome. J. Rheumatol. Suppl. 2000;61:15–21. - PubMed
-
- Sullivan D.A. Sex hormones and Sjogren’s syndrome. J. Rheumatol. Suppl. 1997;50:17–32. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
