

Loop extrusion mediates physiological Igh locus contraction for RAG scanning
- PMID: 33442057
- PMCID: PMC9037962
- DOI: 10.1038/s41586-020-03121-7
Loop extrusion mediates physiological Igh locus contraction for RAG scanning
Abstract
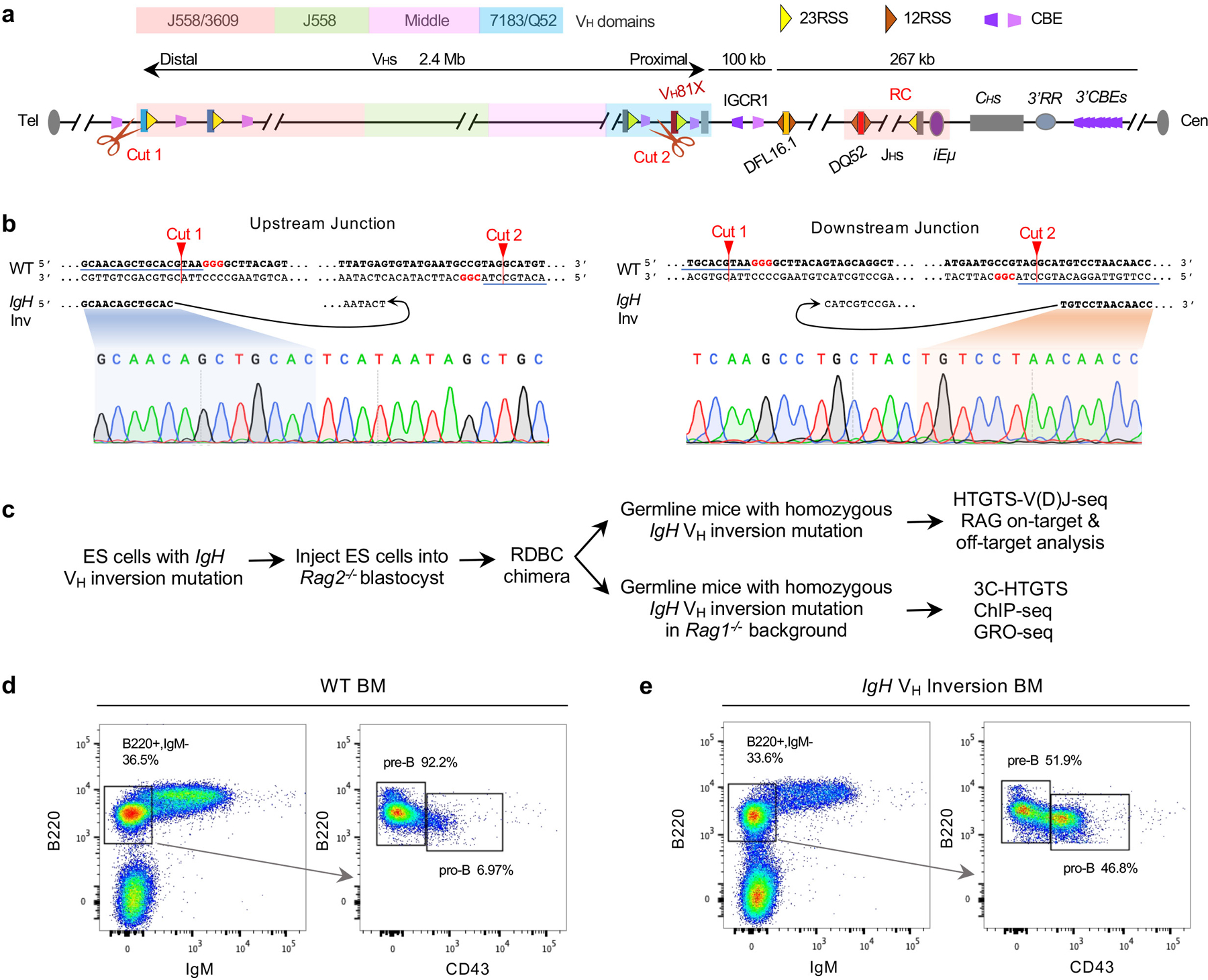
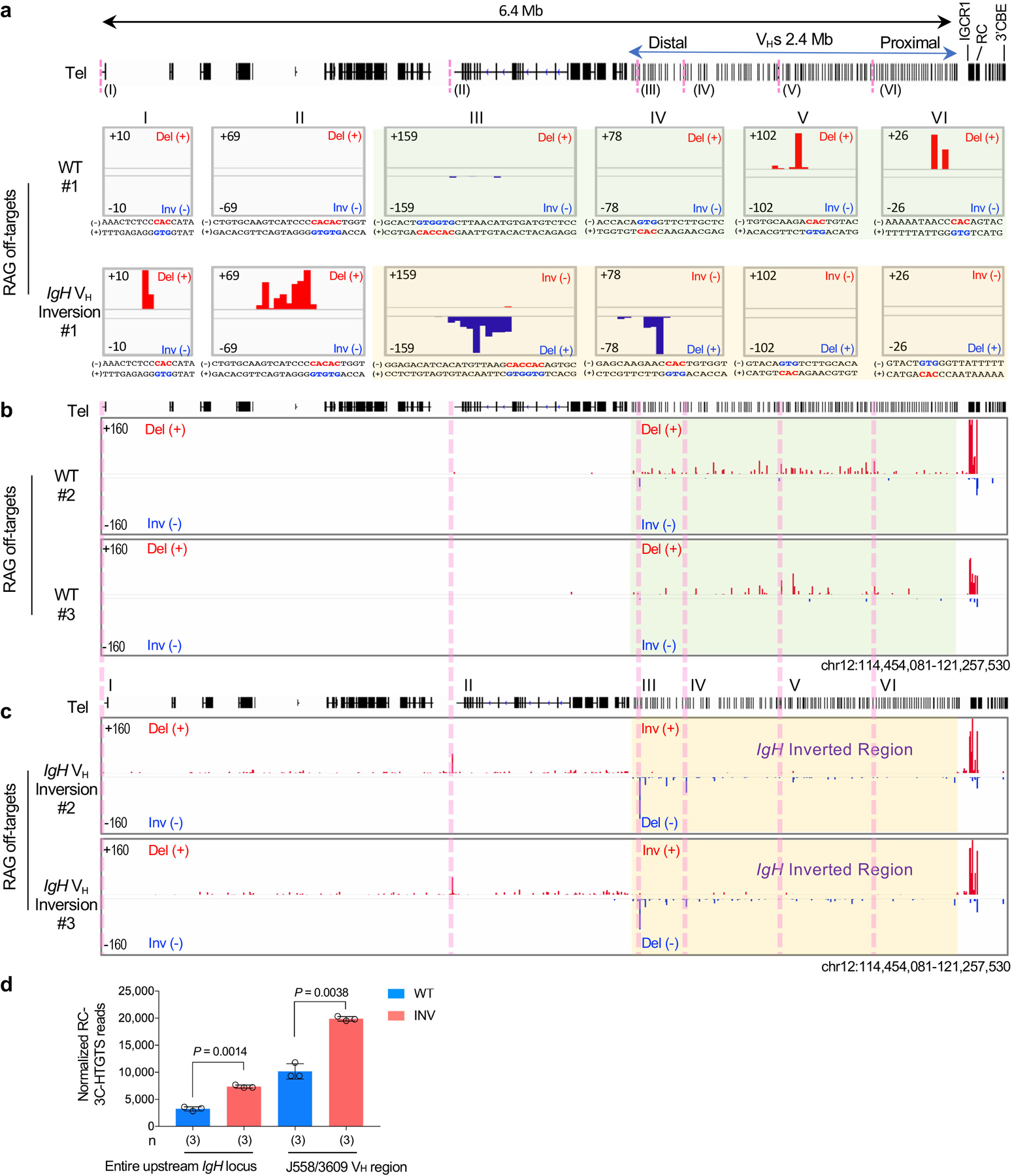
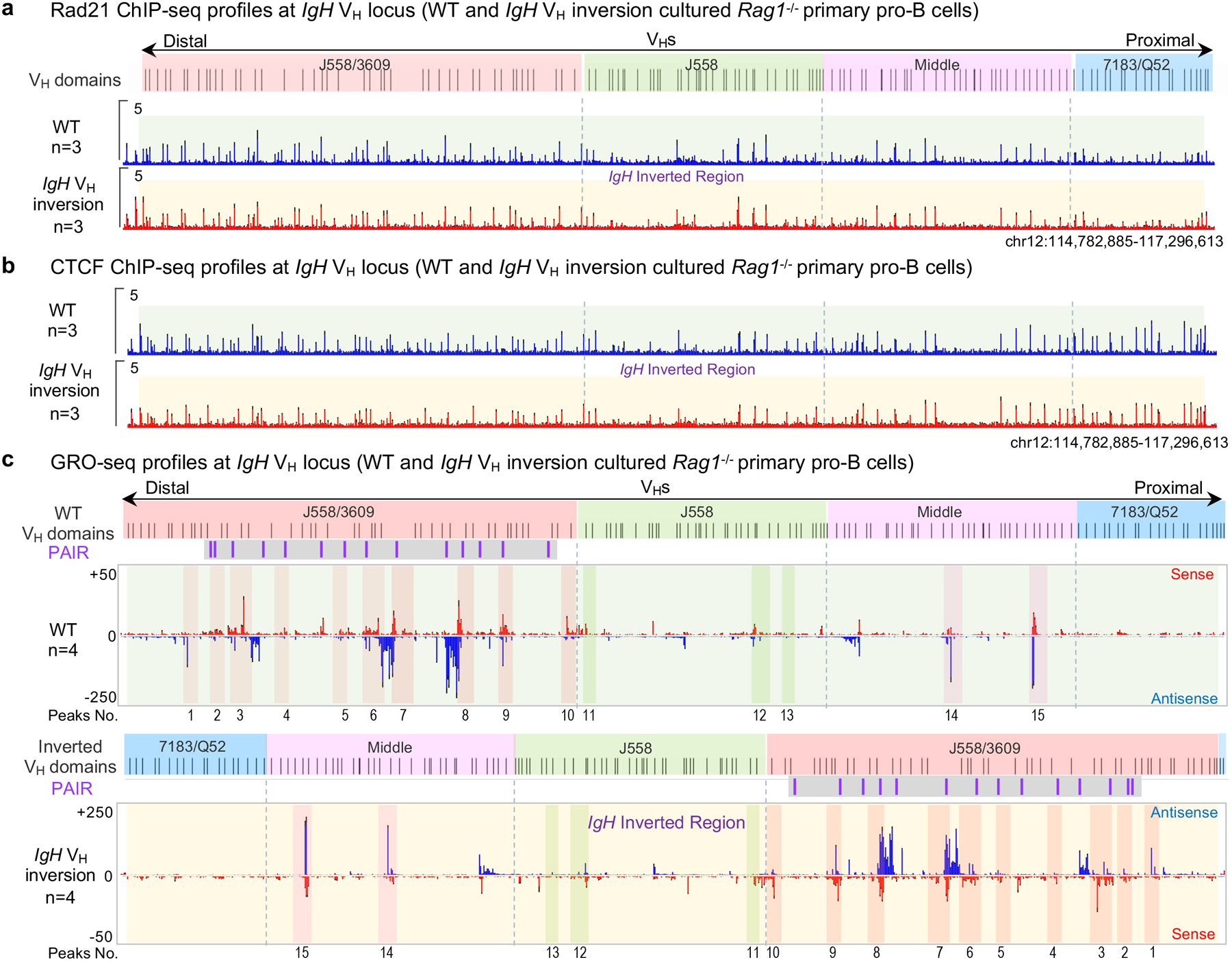
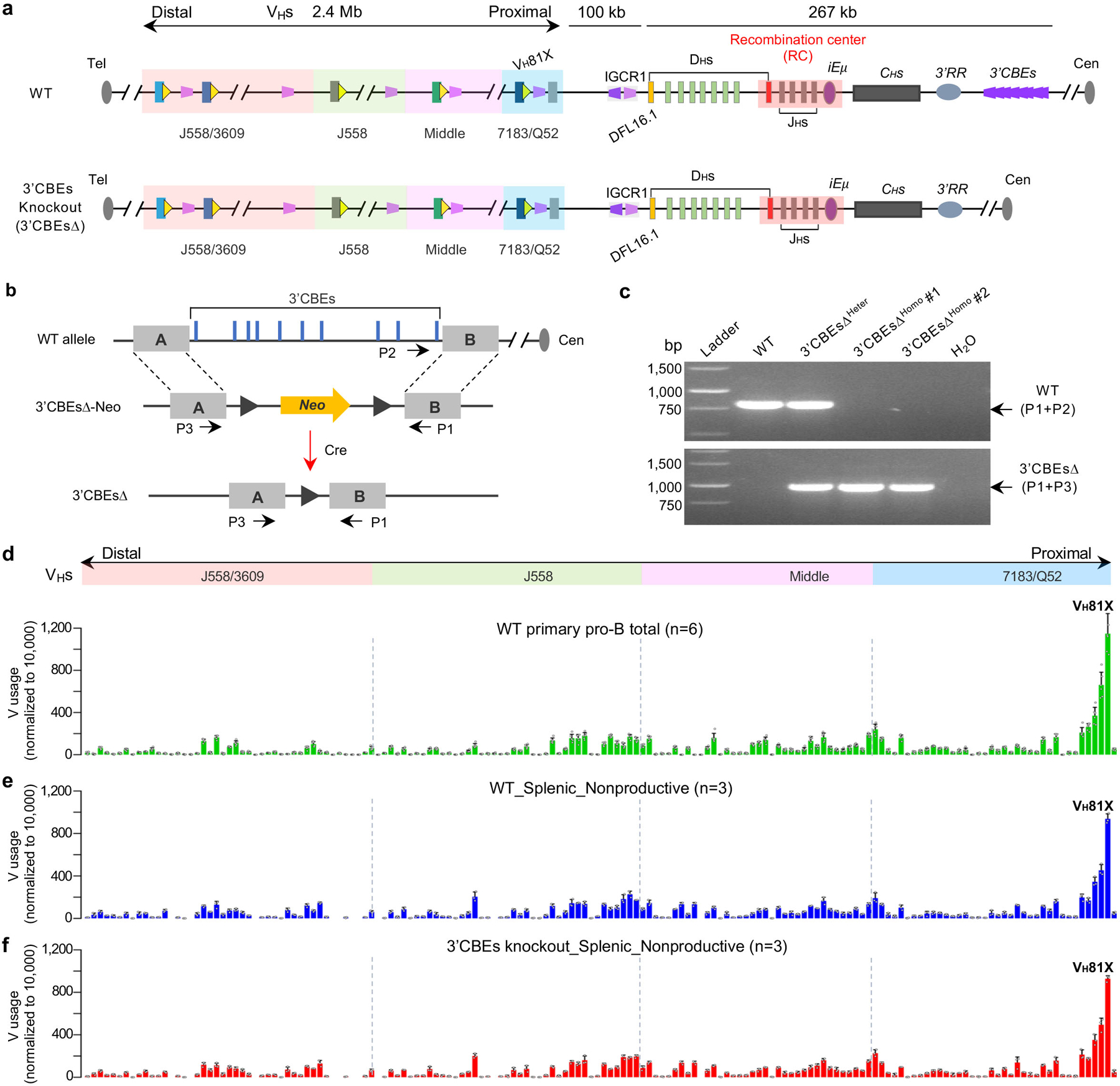
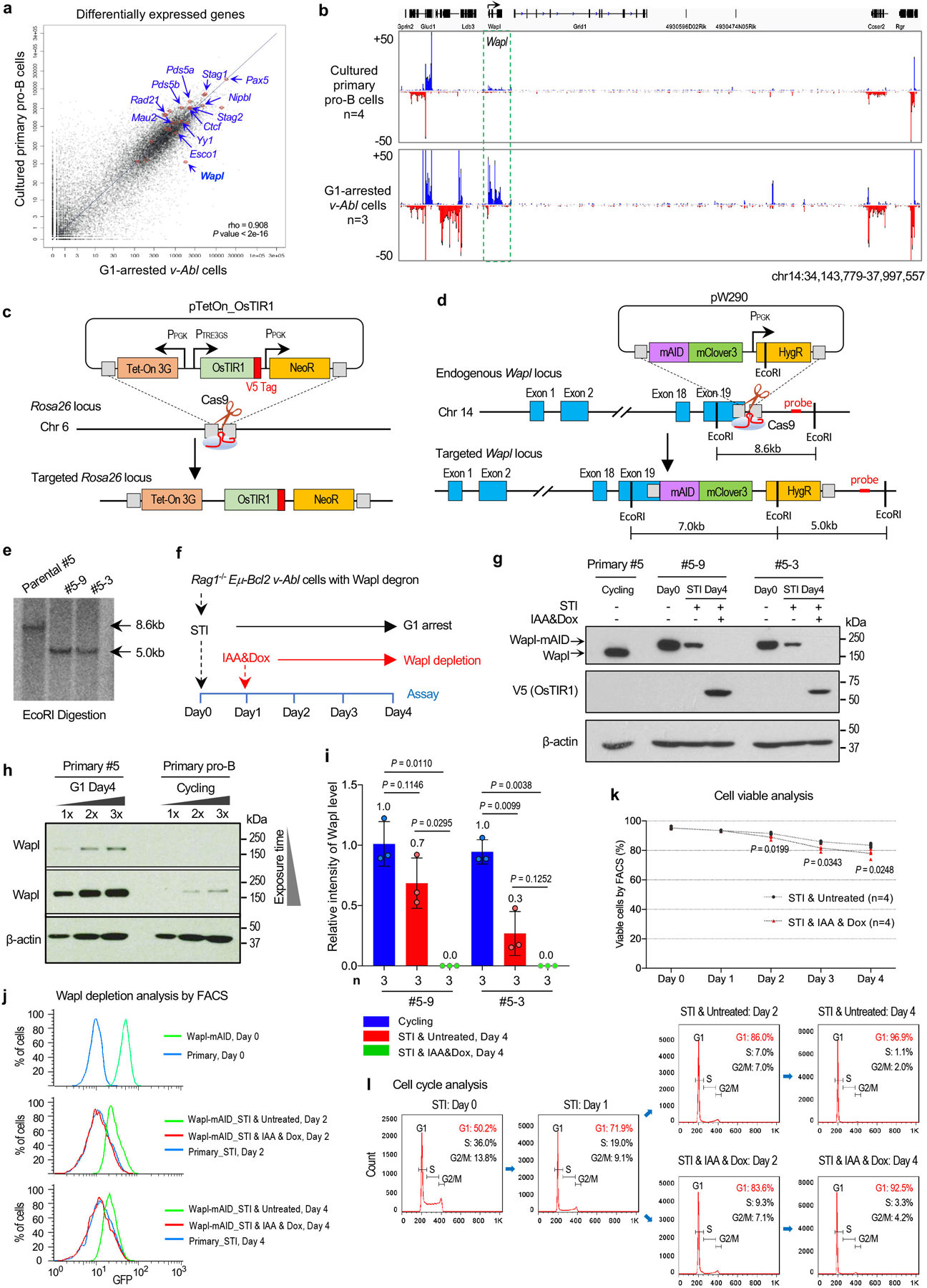
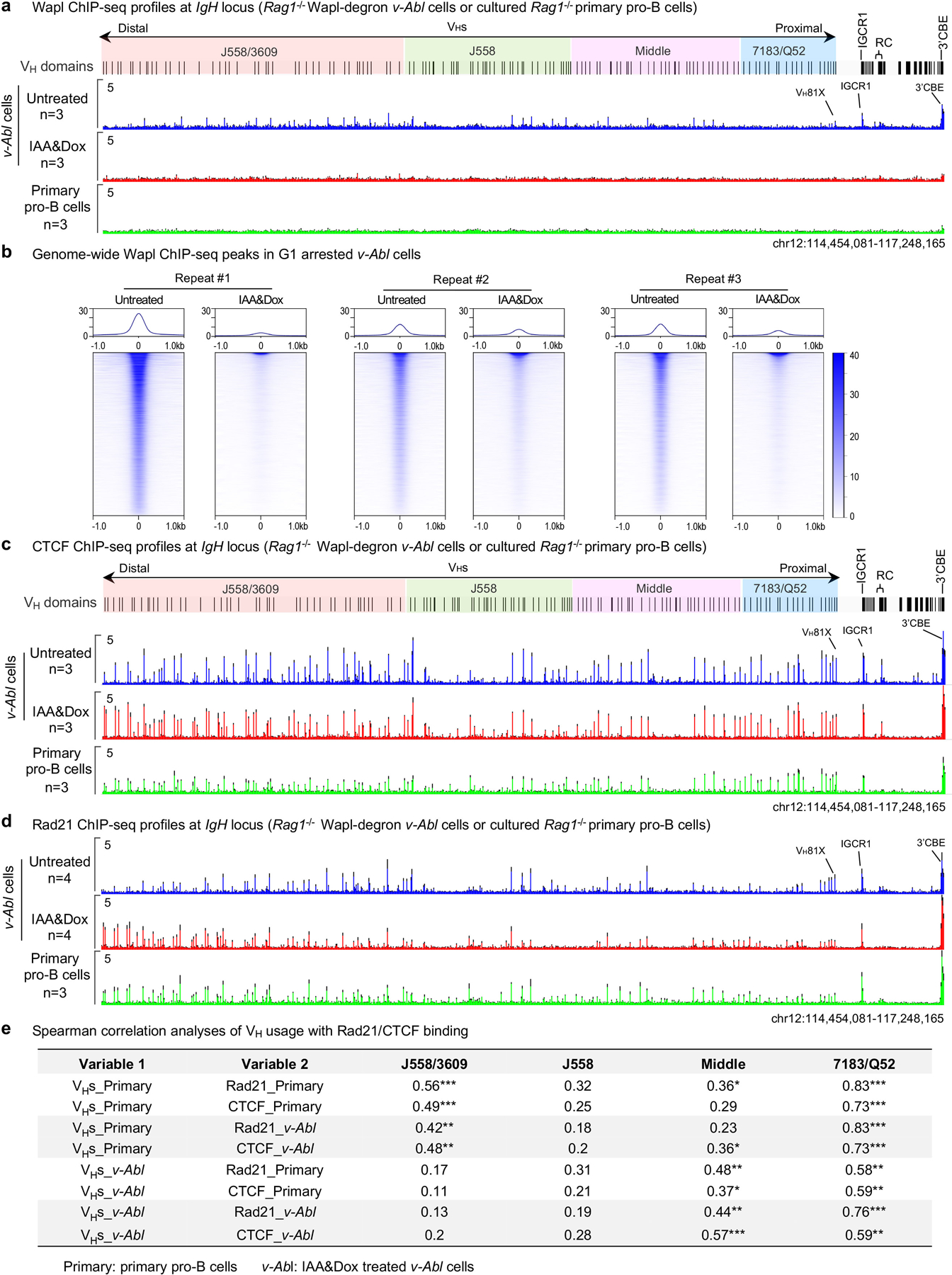
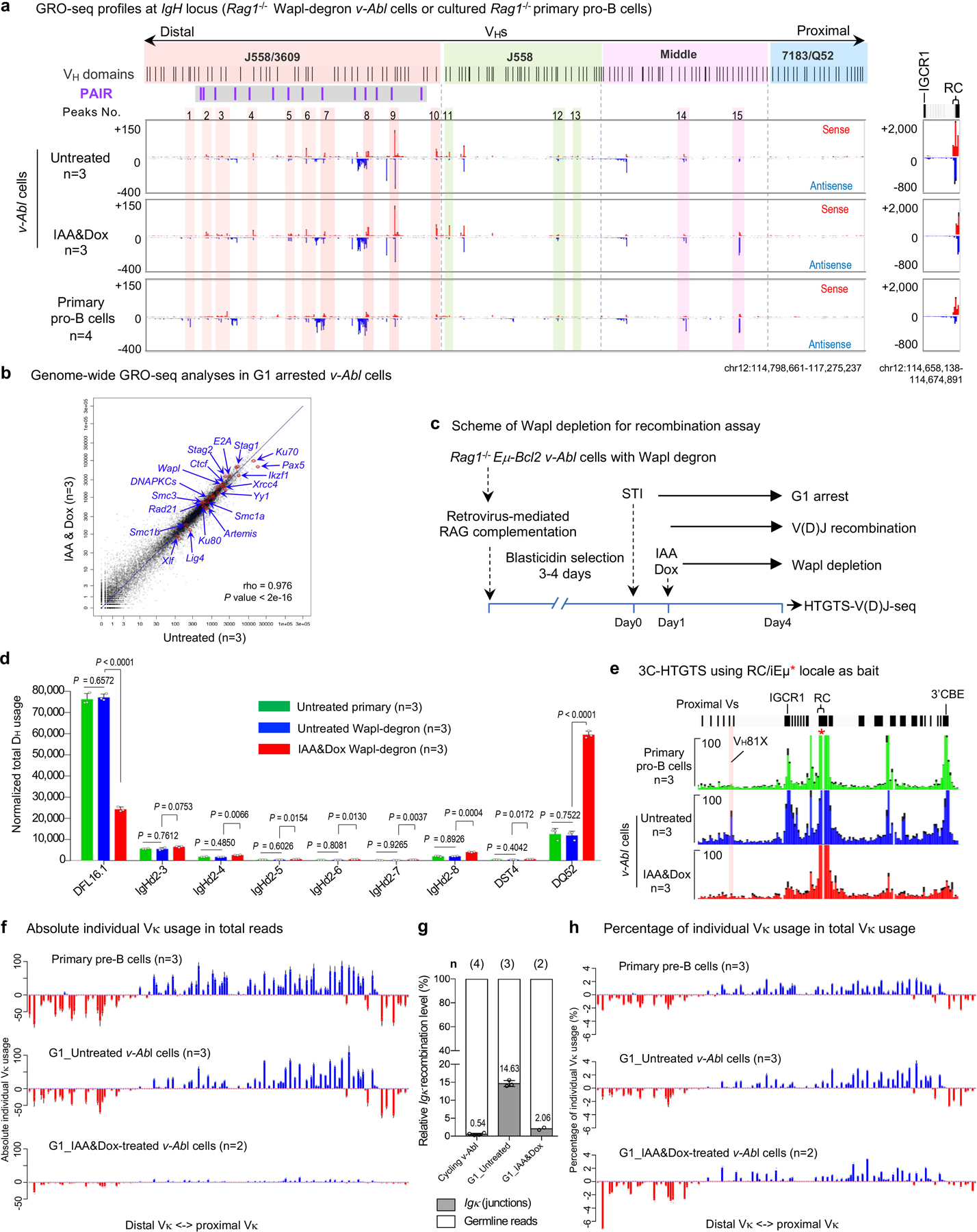
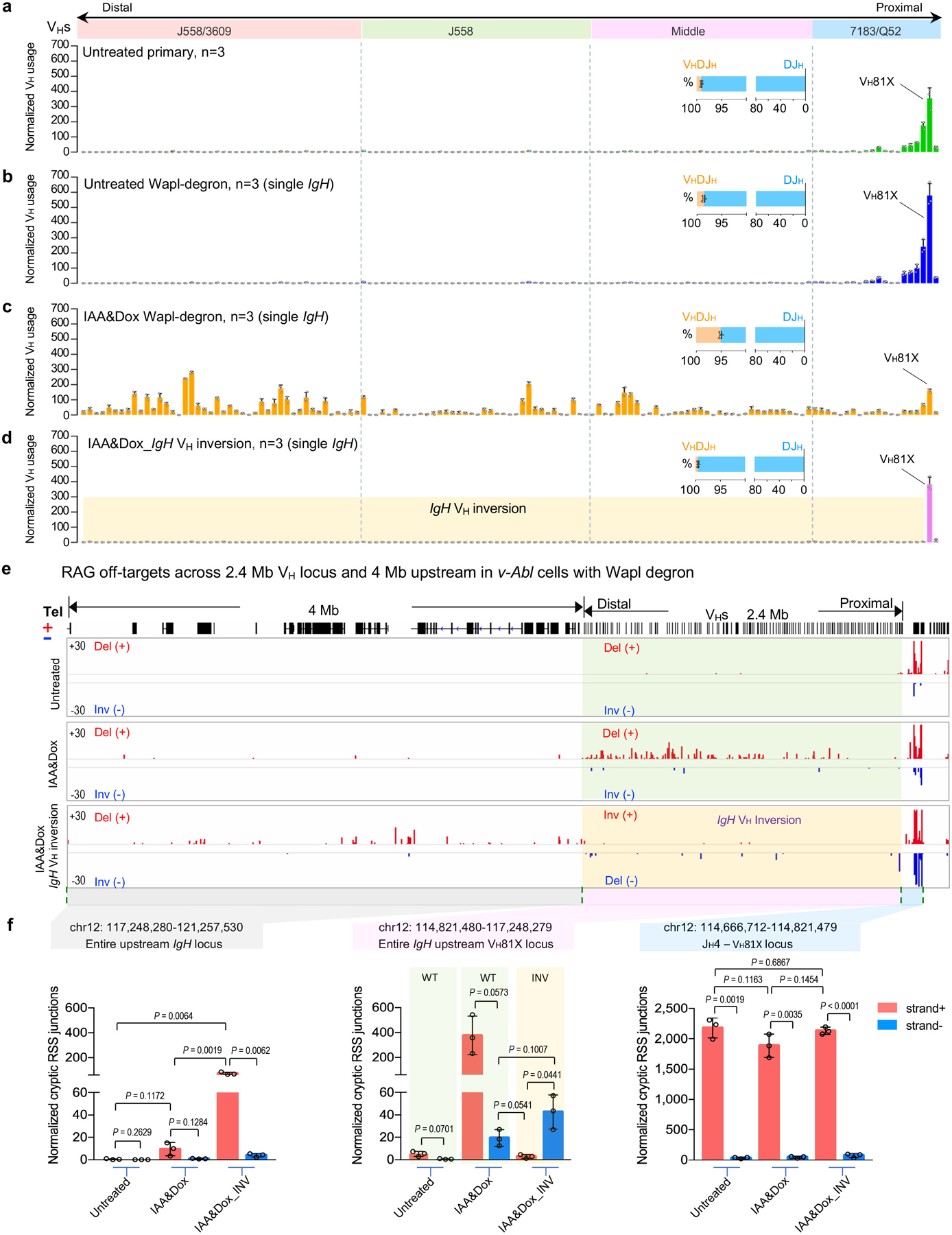
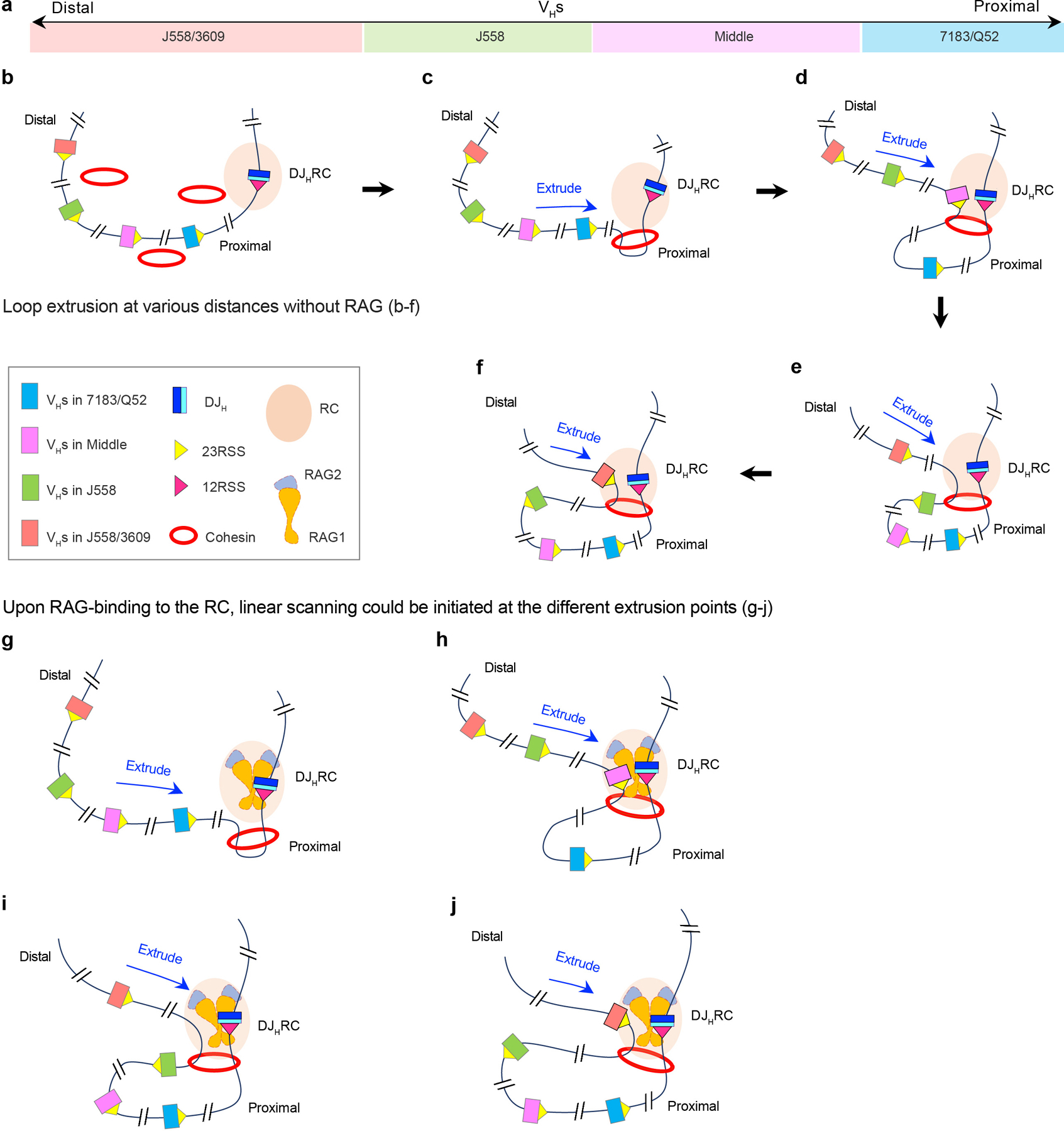
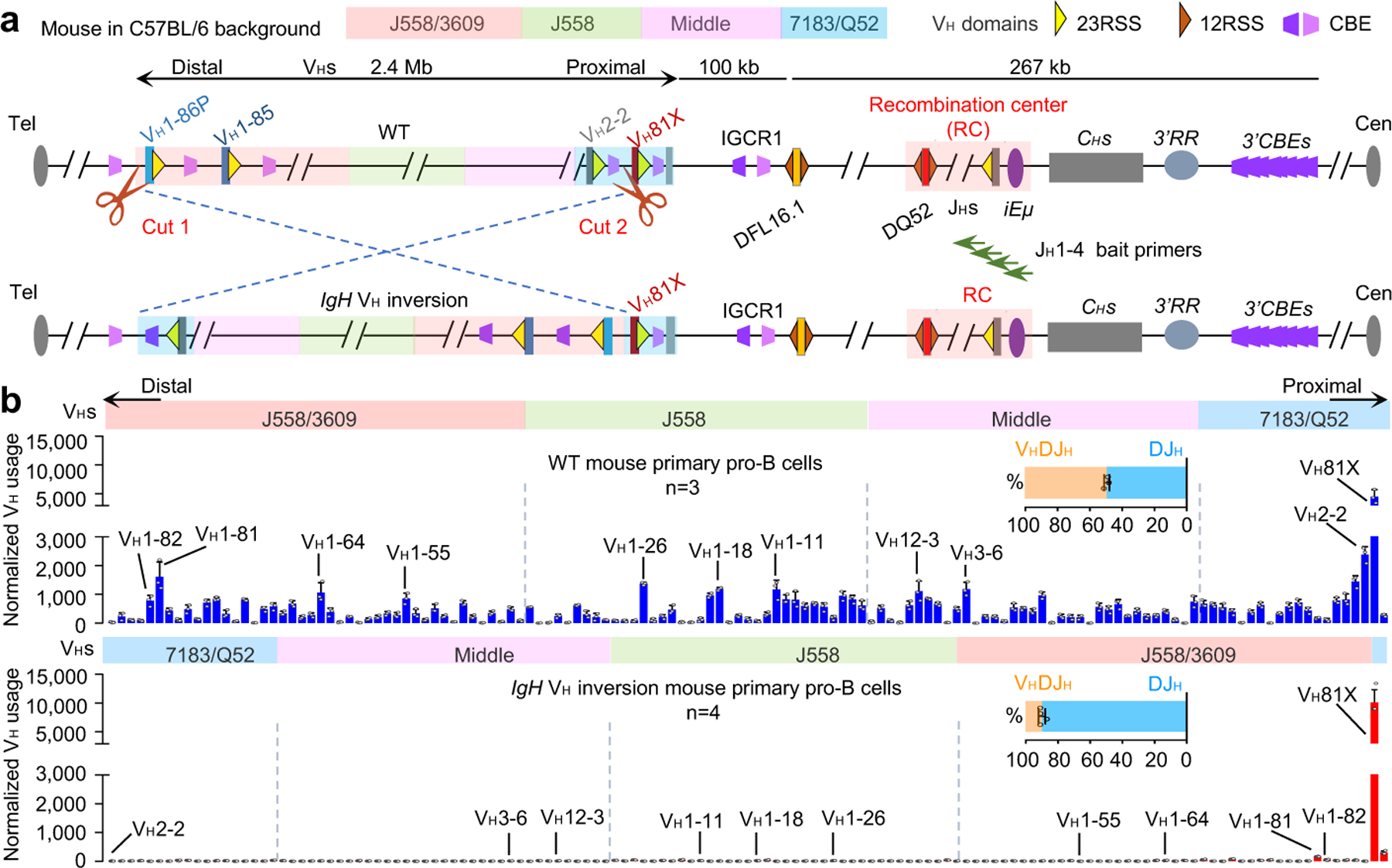
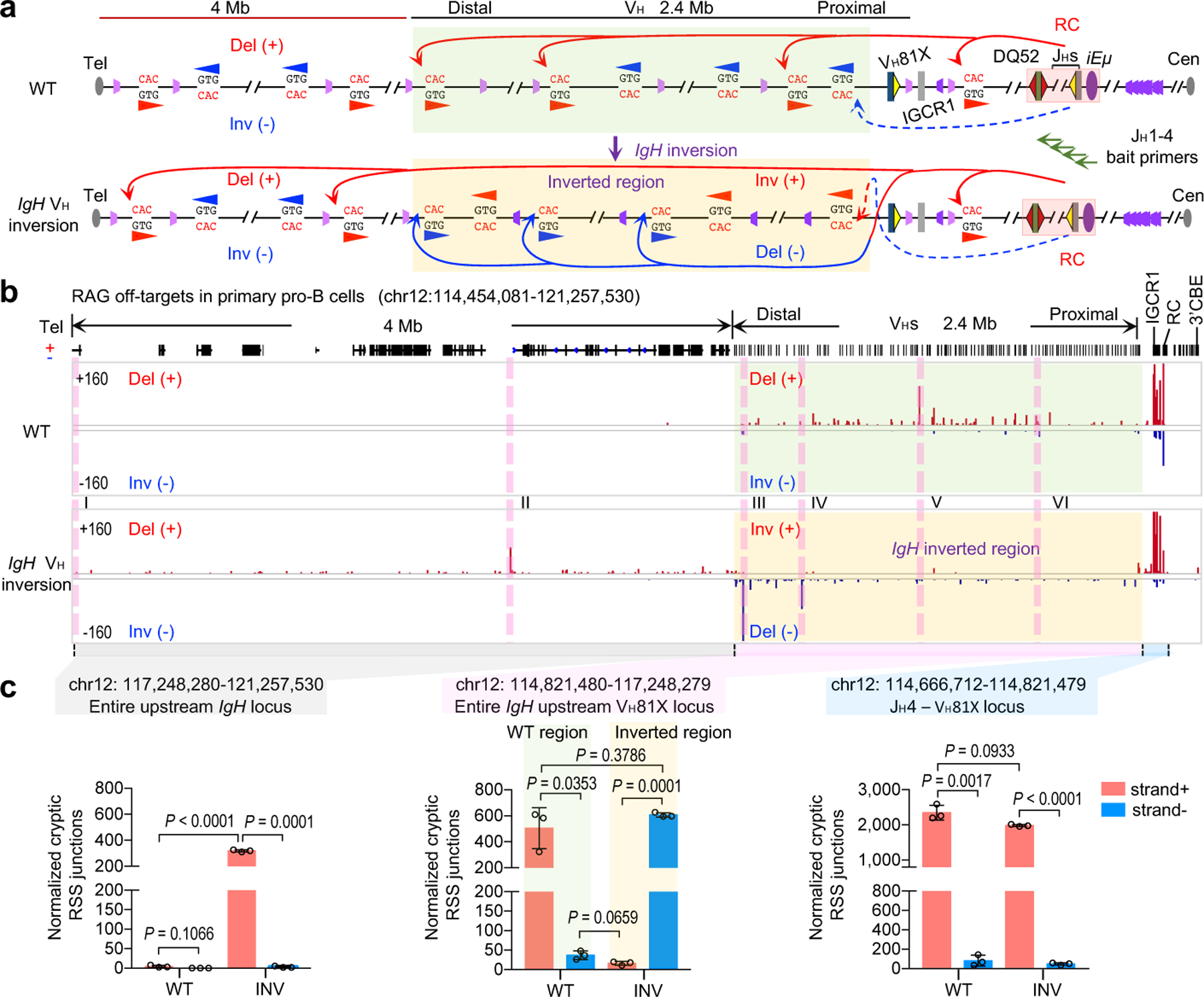
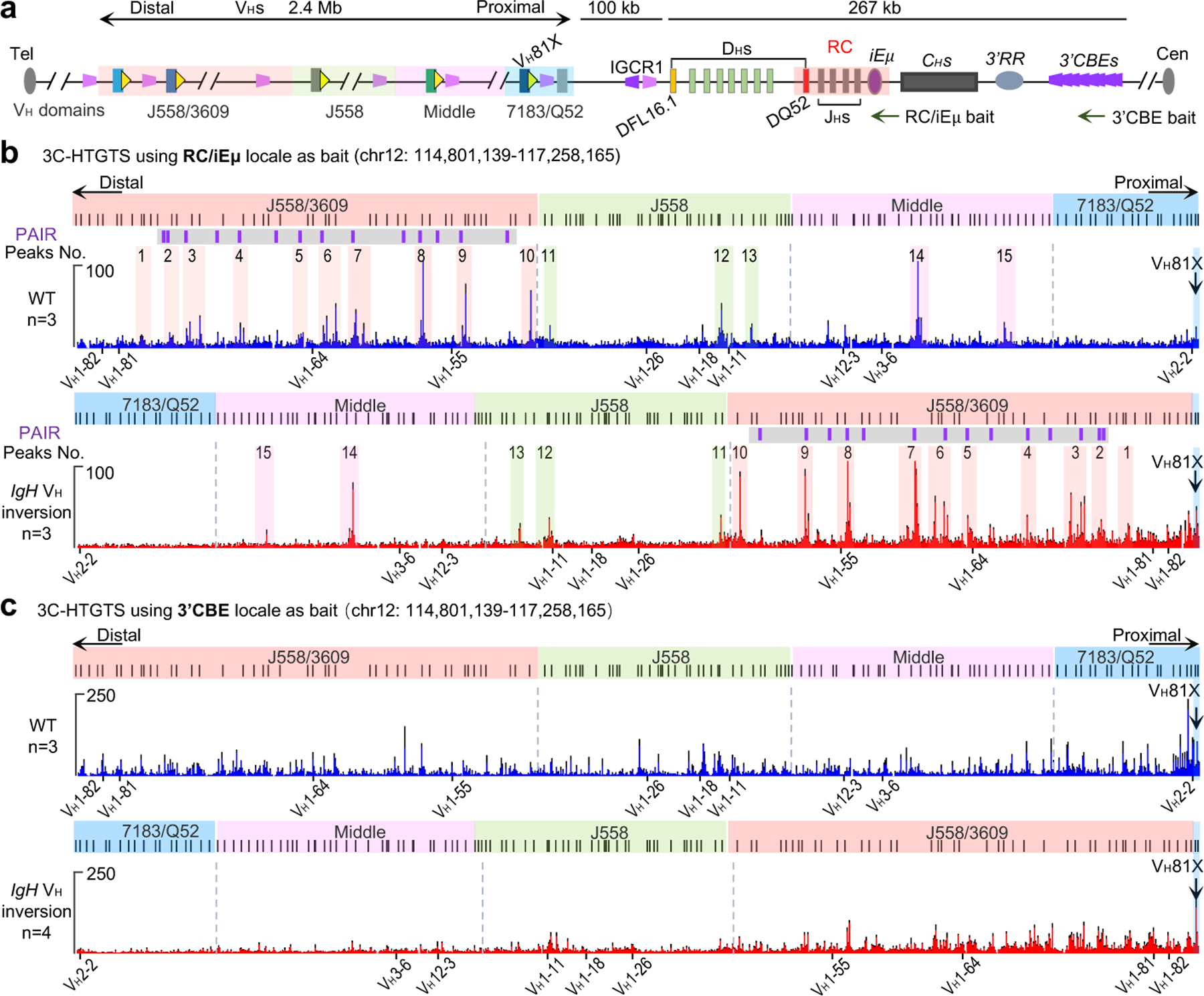
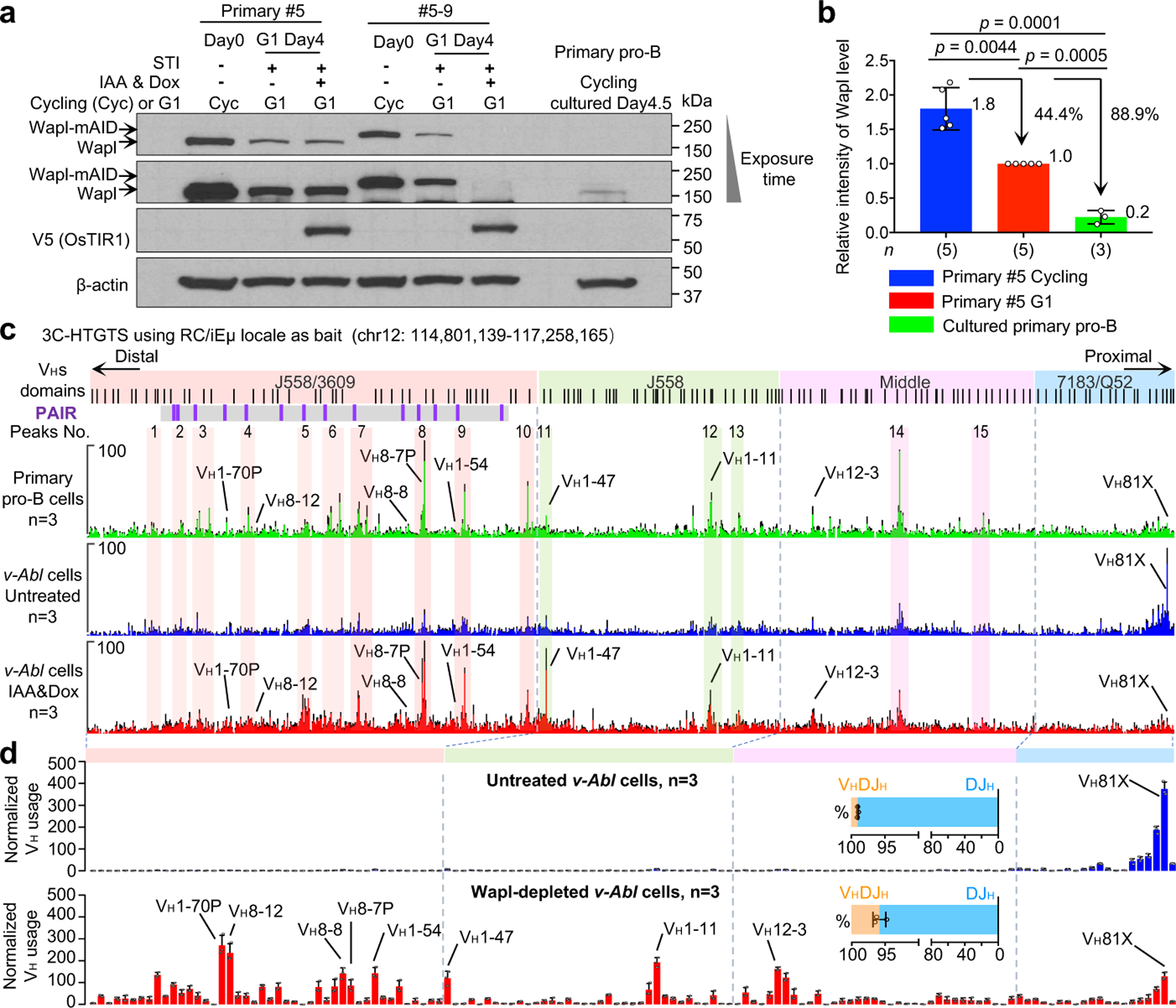
RAG endonuclease initiates Igh V(D)J recombination in progenitor B cells by binding a JH-recombination signal sequence (RSS) within a recombination centre (RC) and then linearly scanning upstream chromatin, presented by loop extrusion mediated by cohesin, for convergent D-RSSs1,2. The utilization of convergently oriented RSSs and cryptic RSSs is intrinsic to long-range RAG scanning3. Scanning of RAG from the DJH-RC-RSS to upstream convergent VH-RSSs is impeded by D-proximal CTCF-binding elements (CBEs)2-5. Primary progenitor B cells undergo a mechanistically undefined contraction of the VH locus that is proposed to provide distal VHs access to the DJH-RC6-9. Here we report that an inversion of the entire 2.4-Mb VH locus in mouse primary progenitor B cells abrogates rearrangement of both VH-RSSs and normally convergent cryptic RSSs, even though locus contraction still occurs. In addition, this inversion activated both the utilization of cryptic VH-RSSs that are normally in opposite orientation and RAG scanning beyond the VH locus through several convergent CBE domains to the telomere. Together, these findings imply that broad deregulation of CBE impediments in primary progenitor B cells promotes RAG scanning of the VH locus mediated by loop extrusion. We further found that the expression of wings apart-like protein homologue (WAPL)10, a cohesin-unloading factor, was low in primary progenitor B cells compared with v-Abl-transformed progenitor B cell lines that lacked contraction and RAG scanning of the VH locus. Correspondingly, depletion of WAPL in v-Abl-transformed lines activated both processes, further implicating loop extrusion in the locus contraction mechanism.
Conflict of interest statement
The authors declare no competing financial interests. F.W.A. is a co-founder of Otoro Biopharmaceuticals.
Figures

References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
