

Genomic Alterations during the In Situ to Invasive Ductal Breast Carcinoma Transition Shaped by the Immune System
- PMID: 33443130
- PMCID: PMC8026652
- DOI: 10.1158/1541-7786.MCR-20-0949
Genomic Alterations during the In Situ to Invasive Ductal Breast Carcinoma Transition Shaped by the Immune System
Abstract
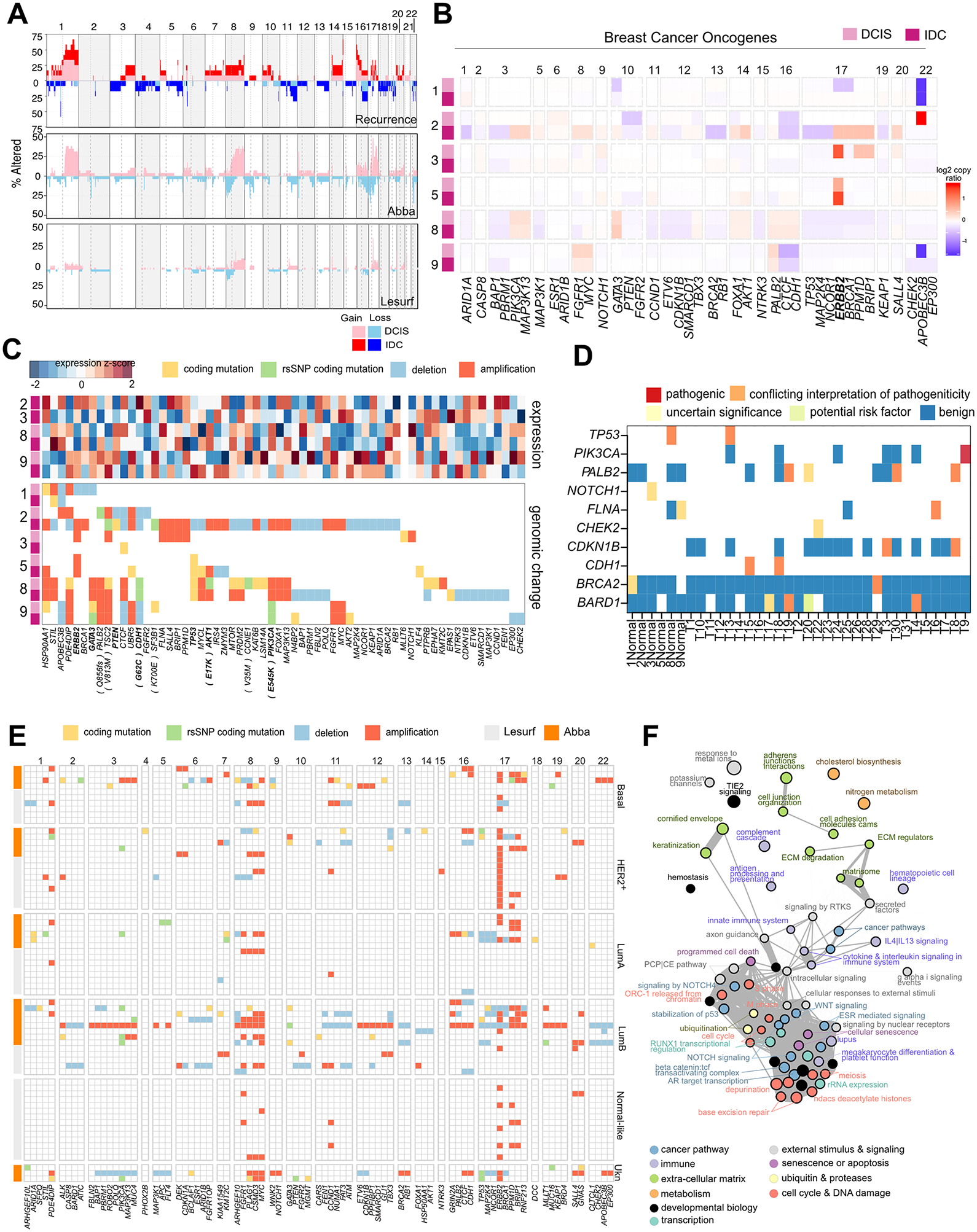
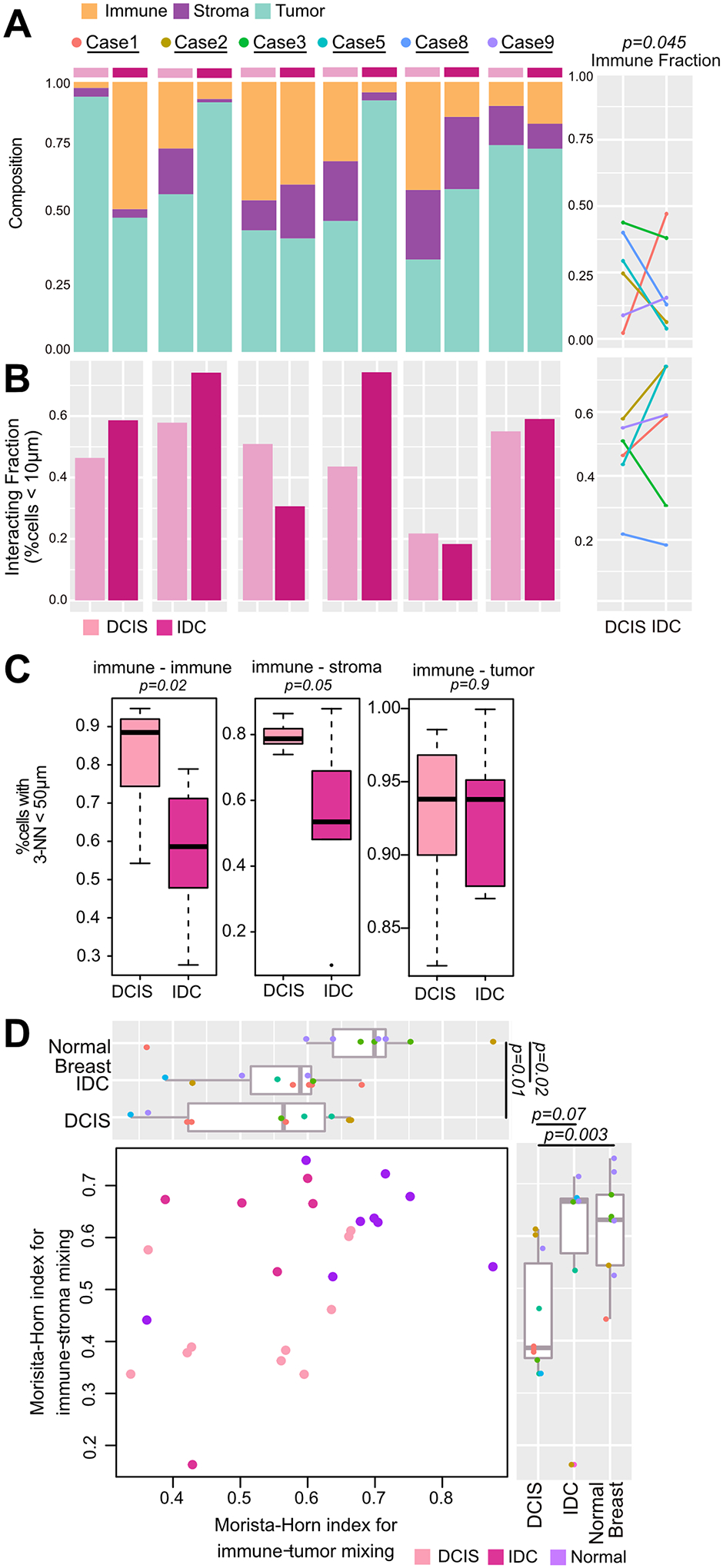
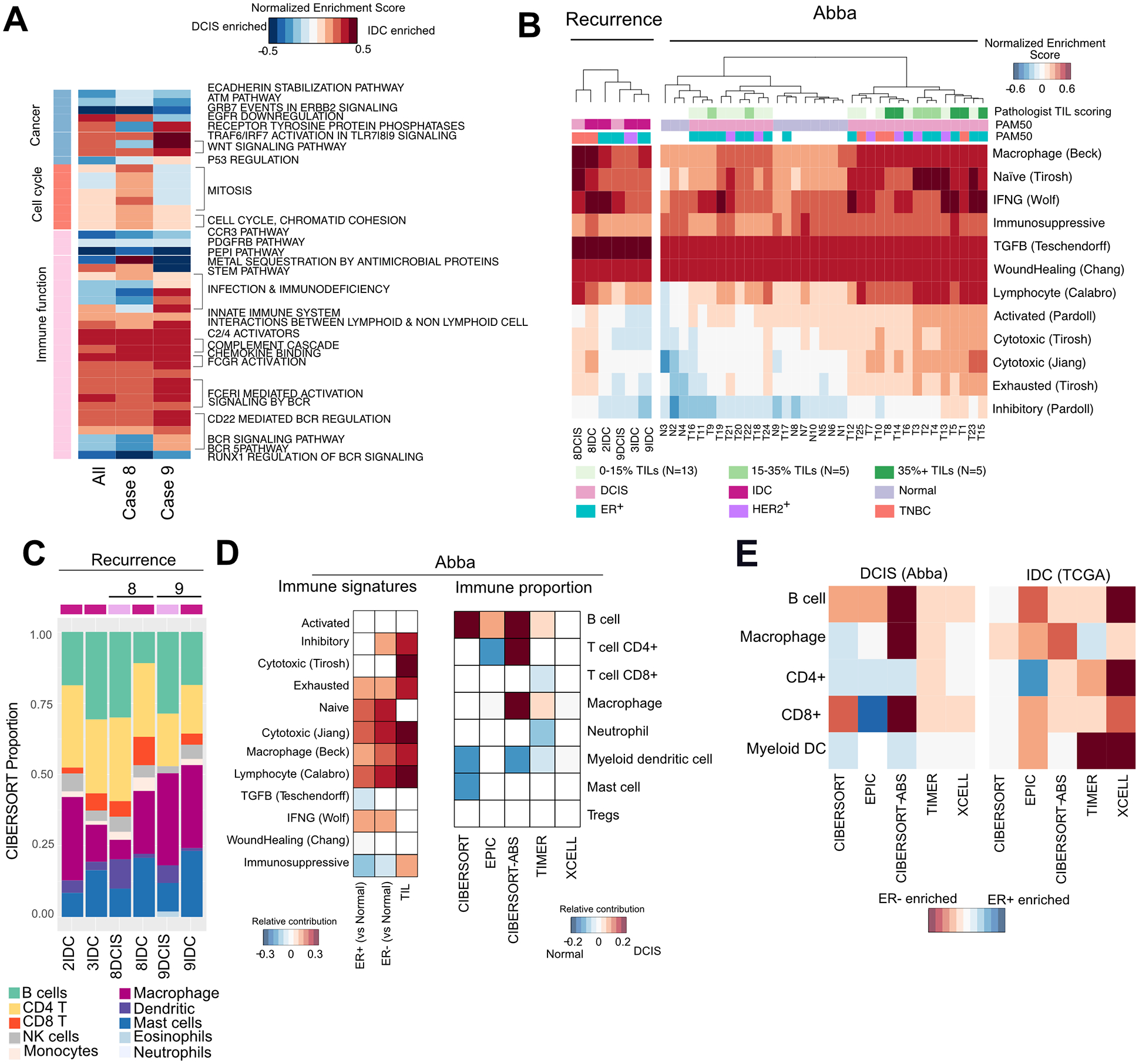
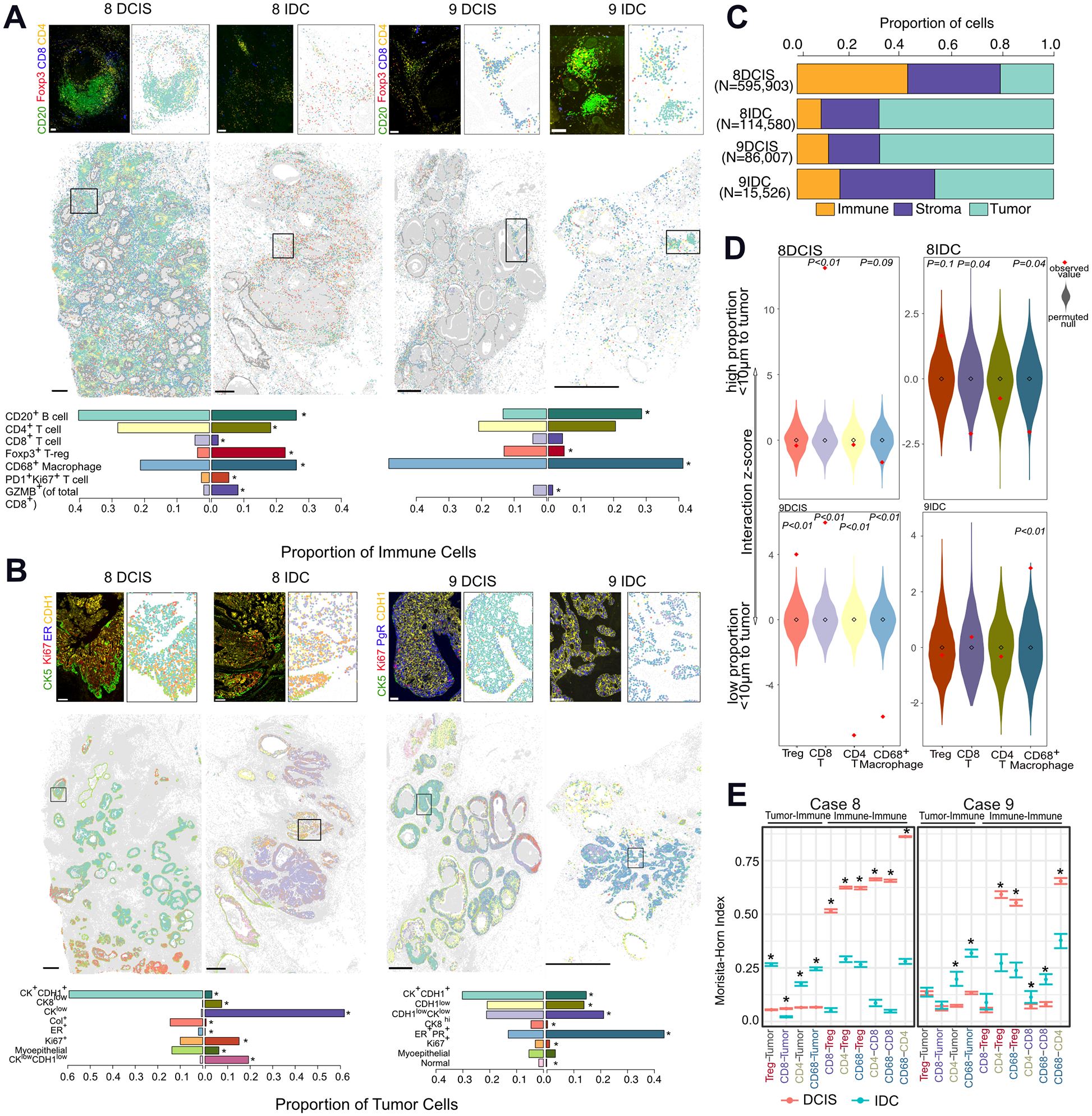
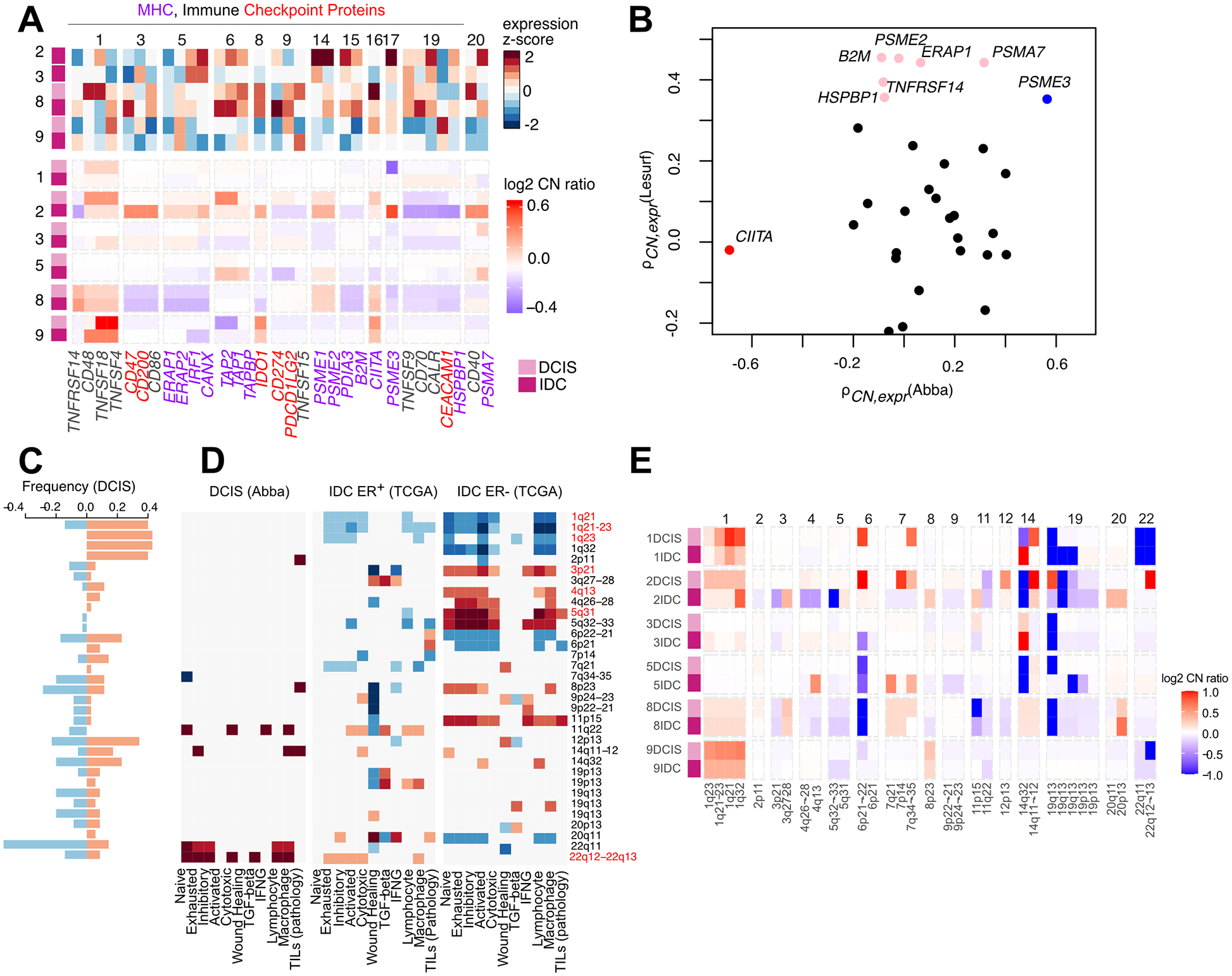
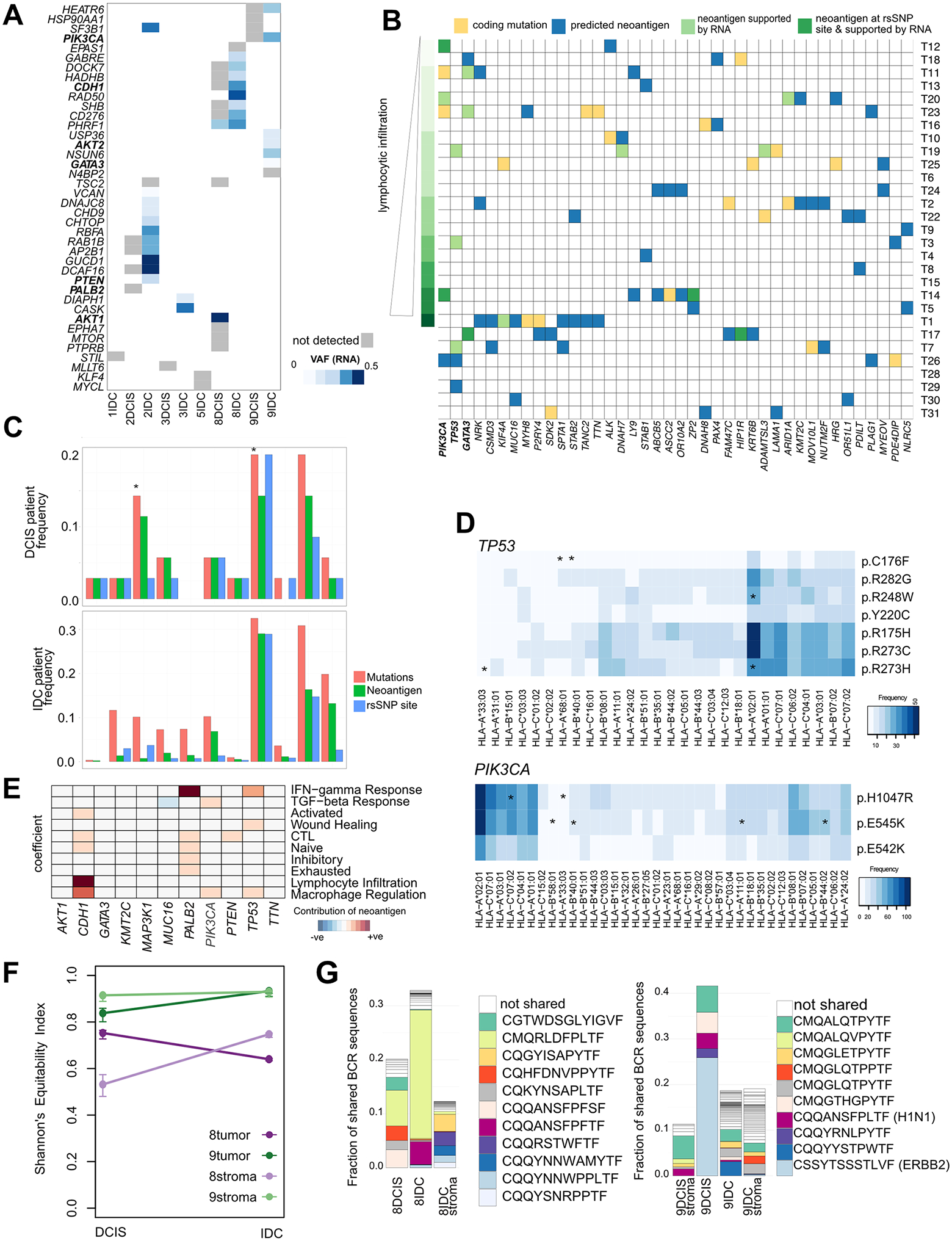
The drivers of ductal carcinoma in situ (DCIS) to invasive ductal carcinoma (IDC) transition are poorly understood. Here, we conducted an integrated genomic, transcriptomic, and whole-slide image analysis to evaluate changes in copy-number profiles, mutational profiles, expression, neoantigen load, and topology in 6 cases of matched pure DCIS and recurrent IDC. We demonstrate through combined copy-number and mutational analysis that recurrent IDC can be genetically related to its pure DCIS despite long latency periods and therapeutic interventions. Immune "hot" and "cold" tumors can arise as early as DCIS and are subtype-specific. Topologic analysis showed a similar degree of pan-leukocyte-tumor mixing in both DCIS and IDC but differ when assessing specific immune subpopulations such as CD4 T cells and CD68 macrophages. Tumor-specific copy-number aberrations in MHC-I presentation machinery and losses in 3p, 4q, and 5p are associated with differences in immune signaling in estrogen receptor (ER)-negative IDC. Common oncogenic hotspot mutations in genes including TP53 and PIK3CA are predicted to be neoantigens yet are paradoxically conserved during the DCIS-to-IDC transition, and are associated with differences in immune signaling. We highlight both tumor and immune-specific changes in the transition of pure DCIS to IDC, including genetic changes in tumor cells that may have a role in modulating immune function and assist in immune escape, driving the transition to IDC. IMPLICATIONS: We demonstrate that the in situ to IDC evolutionary bottleneck is shaped by both tumor and immune cells.
©2020 American Association for Cancer Research.
Conflict of interest statement
Figures
References
-
- Elshof LE, Schaapveld M, Schmidt MK, Rutgers EJ, van Leeuwen FE, Wesseling J. Subsequent risk of ipsilateral and contralateral invasive breast cancer after treatment for ductal carcinoma in situ: incidence and the effect of radiotherapy in a population-based cohort of 10,090 women. Breast Cancer Res Treat 2016;159:553–63. - PMC - PubMed
-
- Gorringe KL, Hunter SM, Pang J-M, Opeskin K, Hill P, Rowley SM, et al. Copy number analysis of ductal carcinoma in situ with and without recurrence. Modern Pathology 2015;28:1174–84. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
