

Evolutionarily related small viral fusogens hijack distinct but modular actin nucleation pathways to drive cell-cell fusion
- PMID: 33443166
- PMCID: PMC7817207
- DOI: 10.1073/pnas.2007526118
Evolutionarily related small viral fusogens hijack distinct but modular actin nucleation pathways to drive cell-cell fusion
Abstract
Fusion-associated small transmembrane (FAST) proteins are a diverse family of nonstructural viral proteins. Once expressed on the plasma membrane of infected cells, they drive fusion with neighboring cells, increasing viral spread and pathogenicity. Unlike viral fusogens with tall ectodomains that pull two membranes together through conformational changes, FAST proteins have short fusogenic ectodomains that cannot bridge the intermembrane gap between neighboring cells. One orthoreovirus FAST protein, p14, has been shown to hijack the actin cytoskeleton to drive cell-cell fusion, but the actin adaptor-binding motif identified in p14 is not found in any other FAST protein. Here, we report that an evolutionarily divergent FAST protein, p22 from aquareovirus, also hijacks the actin cytoskeleton but does so through different adaptor proteins, Intersectin-1 and Cdc42, that trigger N-WASP-mediated branched actin assembly. We show that despite using different pathways, the cytoplasmic tail of p22 can replace that of p14 to create a potent chimeric fusogen, suggesting they are modular and play similar functional roles. When we directly couple p22 with the parallel filament nucleator formin instead of the branched actin nucleation promoting factor N-WASP, its ability to drive fusion is maintained, suggesting that localized mechanical pressure on the plasma membrane coupled to a membrane-disruptive ectodomain is sufficient to drive cell-cell fusion. This work points to a common biophysical strategy used by FAST proteins to push rather than pull membranes together to drive fusion, one that may be harnessed by other short fusogens responsible for physiological cell-cell fusion.
Keywords: FAST proteins; actin cytoskeleton; cell-cell fusion; reovirus.
Conflict of interest statement
The authors declare no competing interest.
Figures
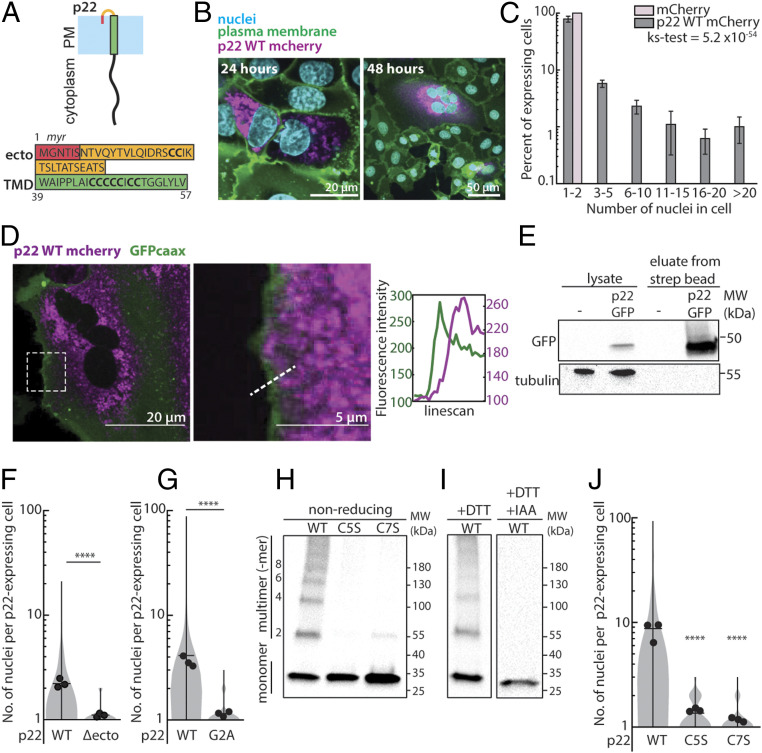
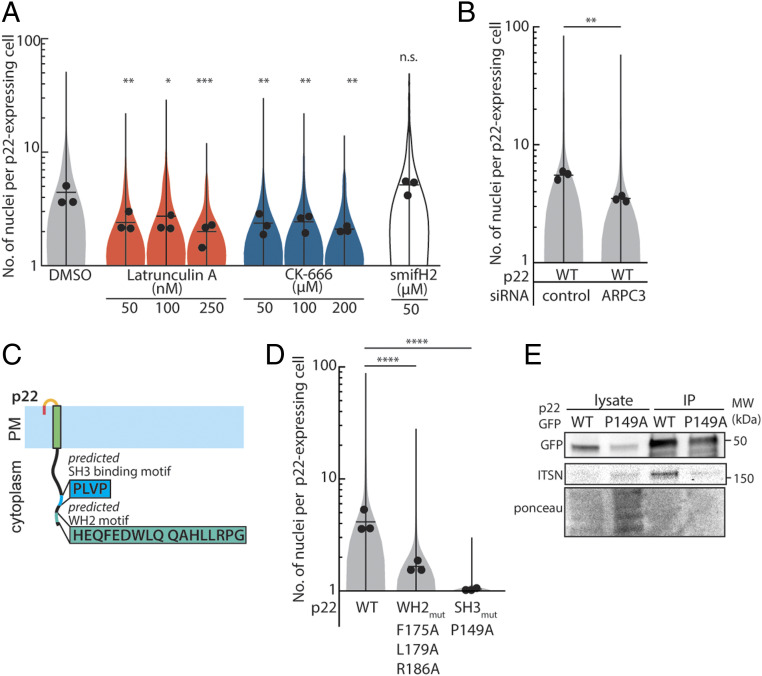
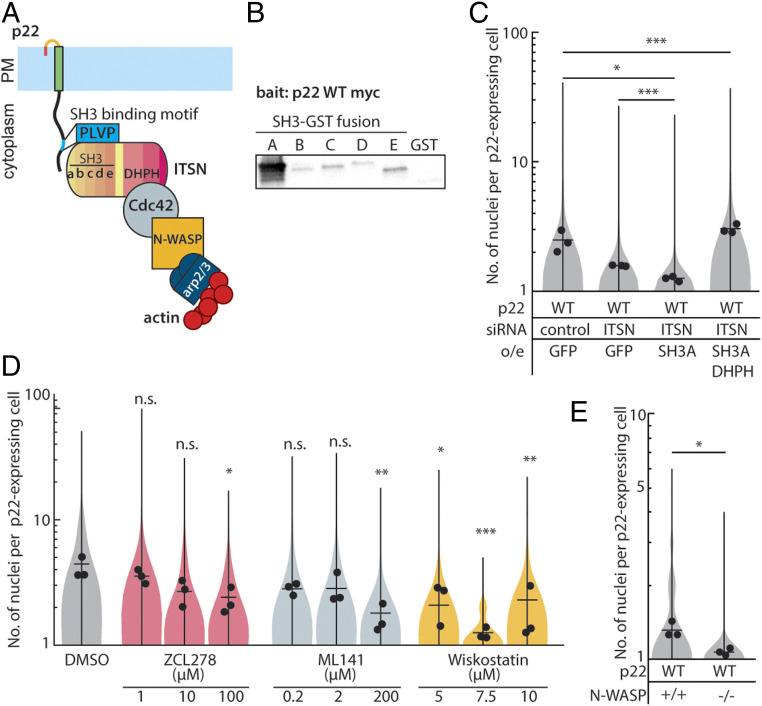
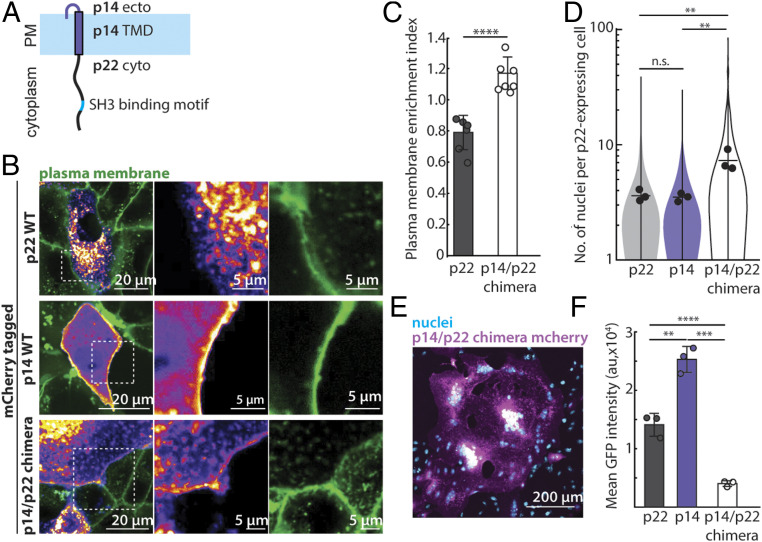
References
-
- Ciechonska M., Duncan R., Reovirus FAST proteins: Virus-encoded cellular fusogens. Trends Microbiol. 22, 715–724 (2014). - PubMed
-
- Duncan R., Fusogenic reoviruses and their fusion-associated small transmembrane (FAST) proteins. Annu. Rev. Virol. 6, 341–363 (2019). - PubMed
-
- Corcoran J. A., et al. , Myristoylation, a protruding loop, and structural plasticity are essential features of a nonenveloped virus fusion peptide motif. J. Biol. Chem. 279, 51386–51394 (2004). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
