

Adipose tissue is a critical regulator of osteoarthritis
- PMID: 33443201
- PMCID: PMC7817130
- DOI: 10.1073/pnas.2021096118
Adipose tissue is a critical regulator of osteoarthritis
Abstract
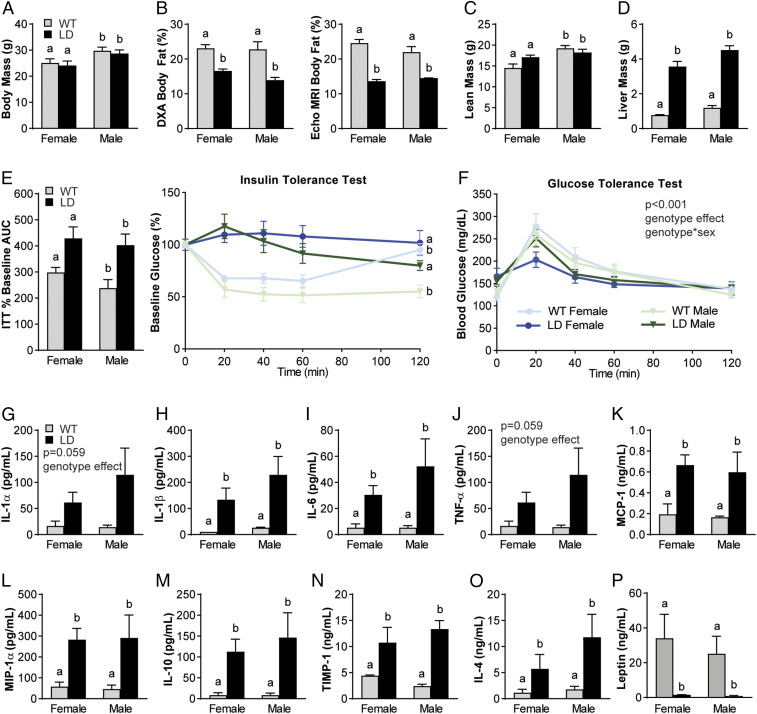
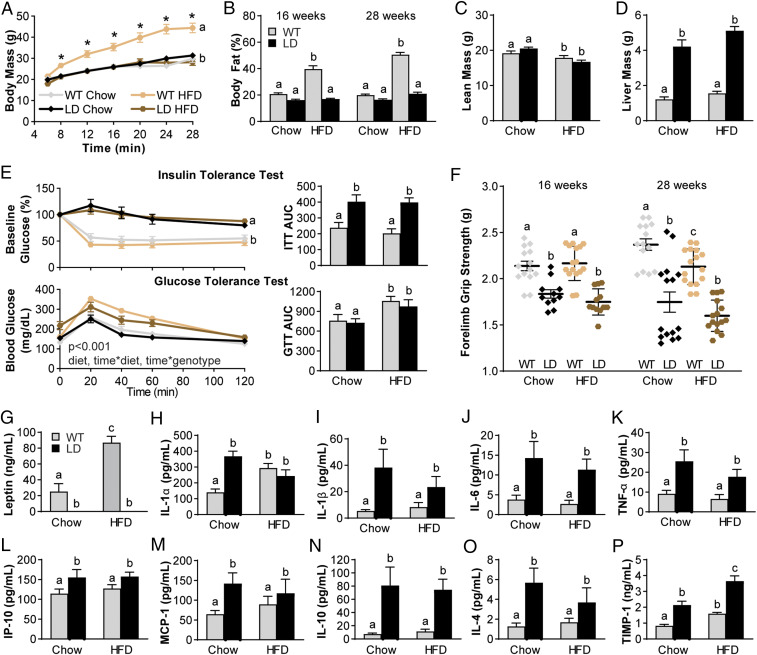
Osteoarthritis (OA), the leading cause of pain and disability worldwide, disproportionally affects individuals with obesity. The mechanisms by which obesity leads to the onset and progression of OA are unclear due to the complex interactions among the metabolic, biomechanical, and inflammatory factors that accompany increased adiposity. We used a murine preclinical model of lipodystrophy (LD) to examine the direct contribution of adipose tissue to OA. Knee joints of LD mice were protected from spontaneous or posttraumatic OA, on either a chow or high-fat diet, despite similar body weight and the presence of systemic inflammation. These findings indicate that adipose tissue itself plays a critical role in the pathophysiology of OA. Susceptibility to posttraumatic OA was reintroduced into LD mice using implantation of a small adipose tissue depot derived from wild-type animals or mouse embryonic fibroblasts that undergo spontaneous adipogenesis, implicating paracrine signaling from fat, rather than body weight, as a mediator of joint degeneration.
Keywords: adipocyte; leptin; muscle weakness; subchondral bone sclerosis; systemic inflammation.
Conflict of interest statement
The authors declare no competing interest.
Figures




Comment in
-
Adipose tissue triggers OA.Nat Rev Rheumatol. 2021 Mar;17(3):130. doi: 10.1038/s41584-021-00579-1. Nat Rev Rheumatol. 2021. PMID: 33504928 No abstract available.
References
-
- Collins K. H., et al. , Relationship between inflammation, the gut microbiota, and metabolic osteoarthritis development: Studies in a rat model. Osteoarthritis Cartilage 23, 1989–1998 (2015). - PubMed
-
- Collins K. H., Reimer R. A., Seerattan R. A., Leonard T. R., Herzog W., Using diet-induced obesity to understand a metabolic subtype of osteoarthritis in rats. Osteoarthritis Cartilage 23, 957–965 (2015). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- S10 OD010707/OD/NIH HHS/United States
- P30 DK056341/DK/NIDDK NIH HHS/United States
- R01 AG015768/AG/NIA NIH HHS/United States
- R61 AR076820/AR/NIAMS NIH HHS/United States
- P30 DK020579/DK/NIDDK NIH HHS/United States
- R37 AR046523/AR/NIAMS NIH HHS/United States
- P30 AR073752/AR/NIAMS NIH HHS/United States
- R01 AG046927/AG/NIA NIH HHS/United States
- P30 AR074992/AR/NIAMS NIH HHS/United States
- T32 DK108742/DK/NIDDK NIH HHS/United States
- P01 AR050245/AR/NIAMS NIH HHS/United States
- R01 DK111389/DK/NIDDK NIH HHS/United States
- T32 DK007120/DK/NIDDK NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
