

Molecular mechanism of the repressive phase of the mammalian circadian clock
- PMID: 33443219
- PMCID: PMC7812753
- DOI: 10.1073/pnas.2021174118
Molecular mechanism of the repressive phase of the mammalian circadian clock
Abstract
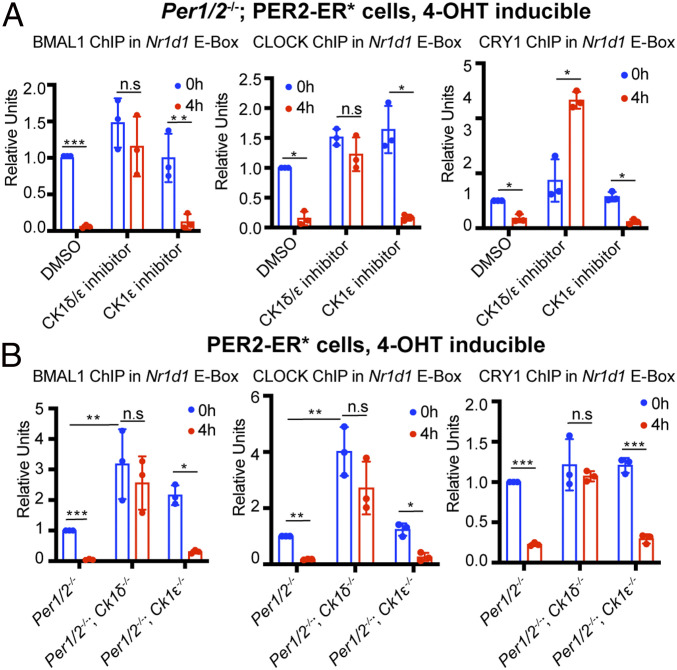
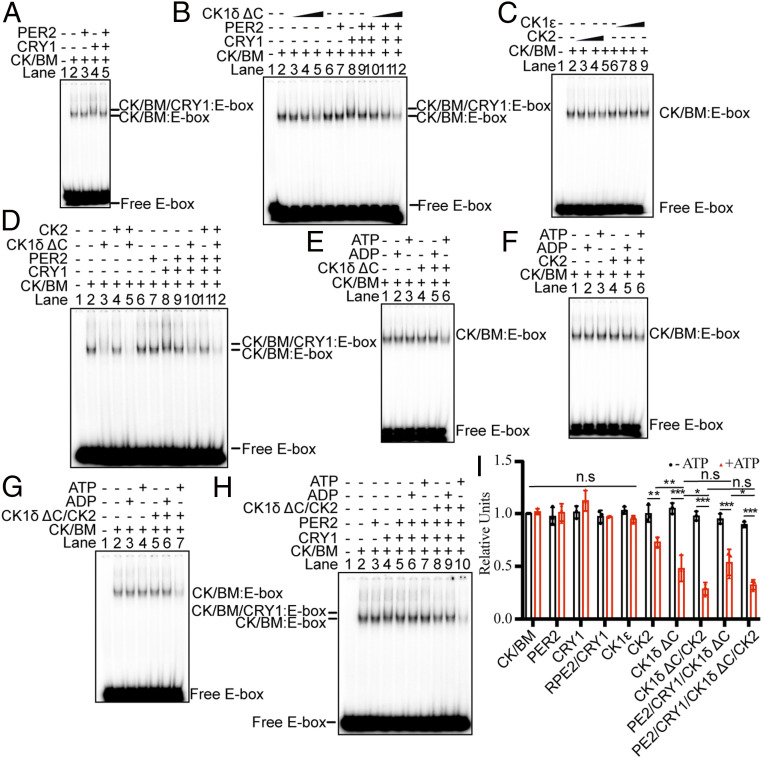
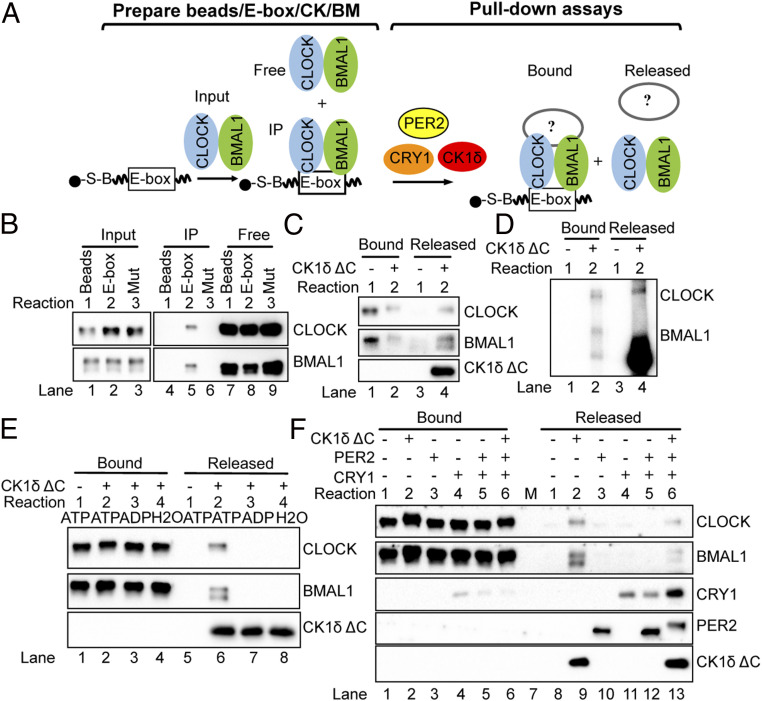
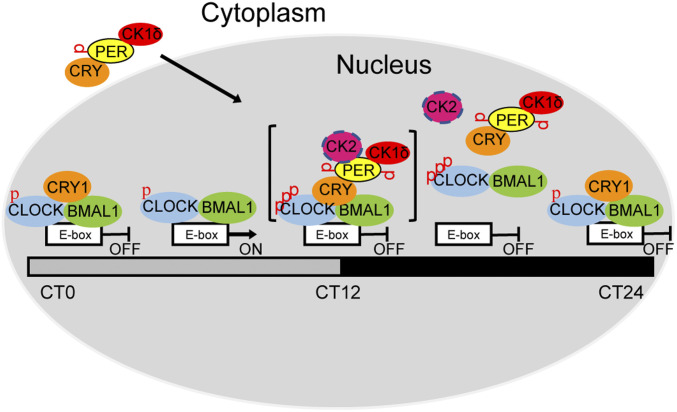
The mammalian circadian clock consists of a transcription-translation feedback loop (TTFL) composed of CLOCK-BMAL1 transcriptional activators and CRY-PER transcriptional repressors. Previous work showed that CRY inhibits CLOCK-BMAL1-activated transcription by a "blocking"-type mechanism and that CRY-PER inhibits CLOCK-BMAL1 by a "displacement"-type mechanism. While the mechanism of CRY-mediated repression was explained by both in vitro and in vivo experiments, the CRY-PER-mediated repression in vivo seemed in conflict with the in vitro data demonstrating PER removes CRY from the CLOCK-BMAL1-E-box complex. Here, we show that CRY-PER participates in the displacement-type repression by recruiting CK1δ to the nucleus and mediating an increased local concentration of CK1δ at CLOCK-BMAL1-bound promoters/enhancers and thus promoting the phosphorylation of CLOCK and dissociation of CLOCK-BMAL1 along with CRY from the E-box. Our findings bring clarity to the role of PER in the dynamic nature of the repressive phase of the TTFL.
Keywords: DNA binding proteins; casein kinase; circadian clock; cryptochrome; period.
Conflict of interest statement
The authors declare no competing interest.
Figures



References
-
- Reppert S. M., Weaver D. R., Coordination of circadian timing in mammals. Nature 418, 935–941 (2002). - PubMed
-
- Hastings M. H., Reddy A. B., Maywood E. S., A clockwork web: Circadian timing in brain and periphery, in health and disease. Nat. Rev. Neurosci. 4, 649–661 (2003). - PubMed
-
- Patke A., Young M. W., Axelrod S., Molecular mechanisms and physiological importance of circadian rhythms. Nat. Rev. Mol. Cell Biol. 21, 67–84 (2020). - PubMed
-
- Sancar A., Mechanisms of DNA repair by photolyase and excision nuclease (Nobel lecture). Angew. Chem. Int. Ed. Engl. 55, 8502–8527 (2016). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
