

Ecd promotes U5 snRNP maturation and Prp8 stability
- PMID: 33444449
- PMCID: PMC7897482
- DOI: 10.1093/nar/gkaa1274
Ecd promotes U5 snRNP maturation and Prp8 stability
Abstract
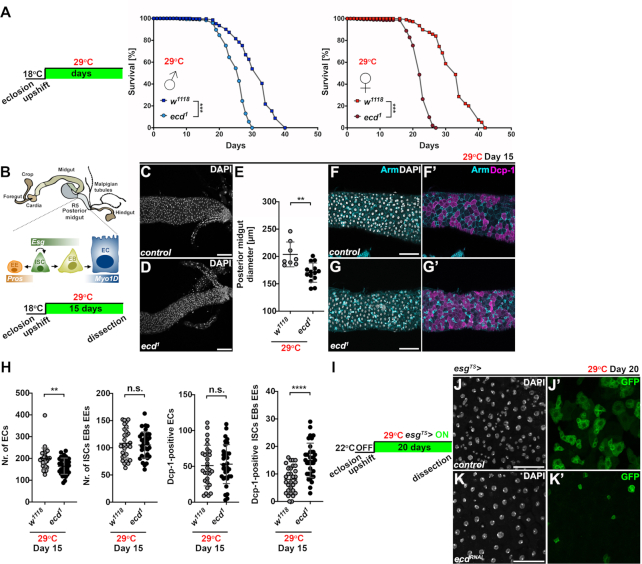
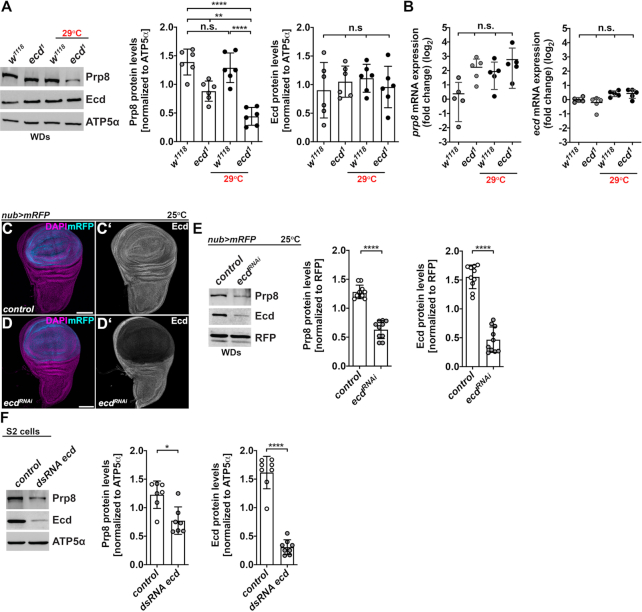
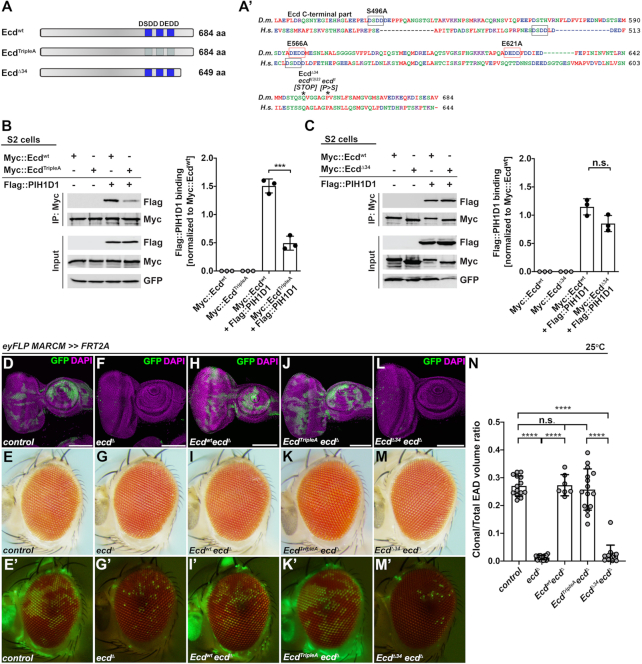
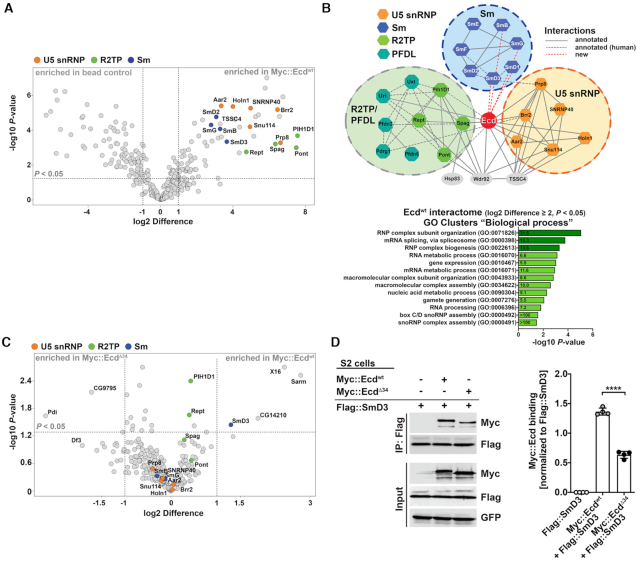
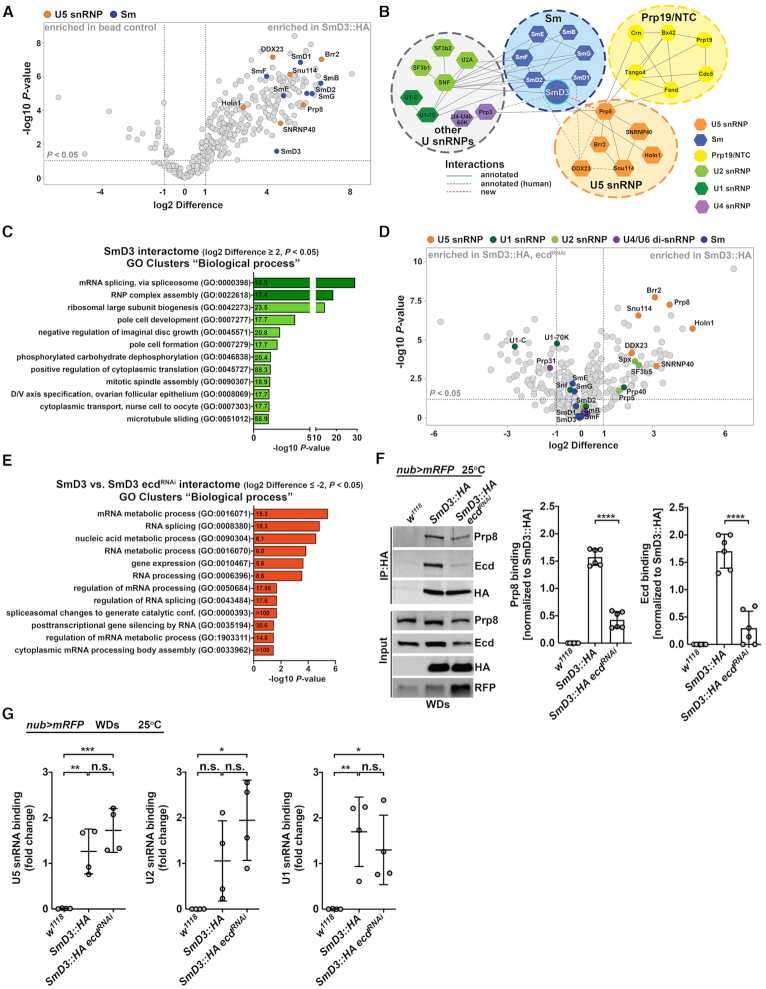
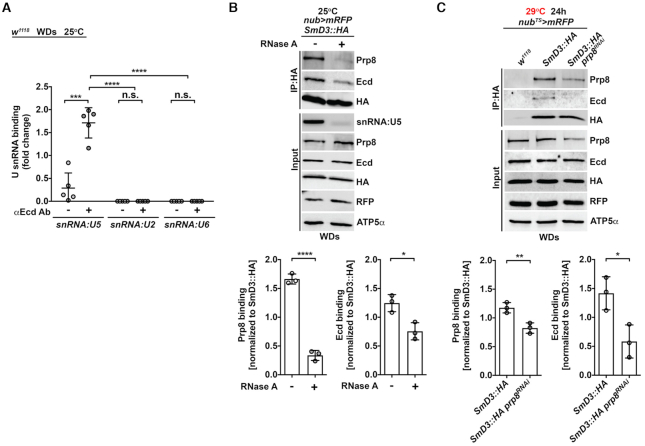
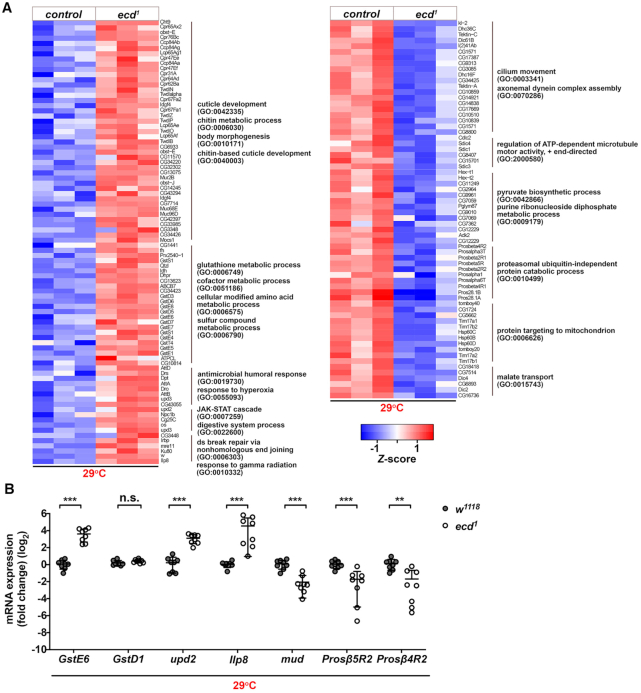
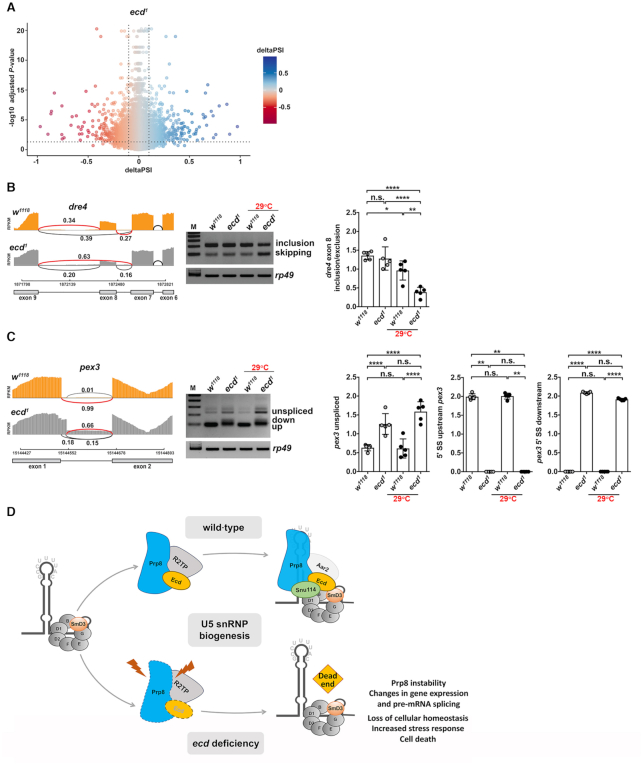
Pre-mRNA splicing catalyzed by the spliceosome represents a critical step in the regulation of gene expression contributing to transcriptome and proteome diversity. The spliceosome consists of five small nuclear ribonucleoprotein particles (snRNPs), the biogenesis of which remains only partially understood. Here we define the evolutionarily conserved protein Ecdysoneless (Ecd) as a critical regulator of U5 snRNP assembly and Prp8 stability. Combining Drosophila genetics with proteomic approaches, we demonstrate the Ecd requirement for the maintenance of adult healthspan and lifespan and identify the Sm ring protein SmD3 as a novel interaction partner of Ecd. We show that the predominant task of Ecd is to deliver Prp8 to the emerging U5 snRNPs in the cytoplasm. Ecd deficiency, on the other hand, leads to reduced Prp8 protein levels and compromised U5 snRNP biogenesis, causing loss of splicing fidelity and transcriptome integrity. Based on our findings, we propose that Ecd chaperones Prp8 to the forming U5 snRNP allowing completion of the cytoplasmic part of the U5 snRNP biogenesis pathway necessary to meet the cellular demand for functional spliceosomes.
© The Author(s) 2021. Published by Oxford University Press on behalf of Nucleic Acids Research.
Figures
References
-
- Fischer U., Englbrecht C., Chari A.. Biogenesis of spliceosomal small nuclear ribonucleoproteins. Wiley Interdiscip. Rev. RNA. 2011; 2:718–731. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
