

Kappa opioid receptor activation in the amygdala disinhibits CRF neurons to generate pain-like behaviors
- PMID: 33444637
- PMCID: PMC7887082
- DOI: 10.1016/j.neuropharm.2021.108456
Kappa opioid receptor activation in the amygdala disinhibits CRF neurons to generate pain-like behaviors
Abstract
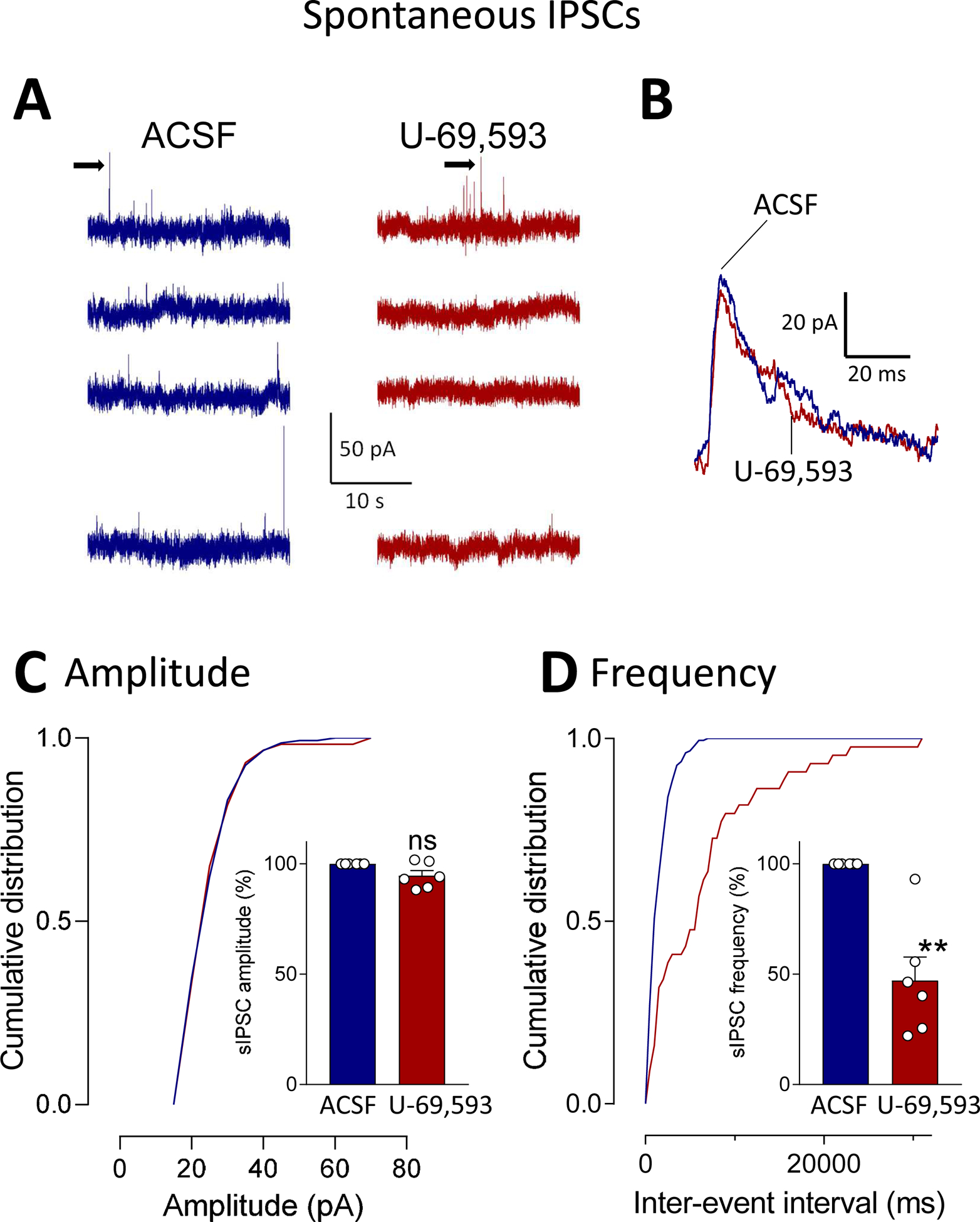
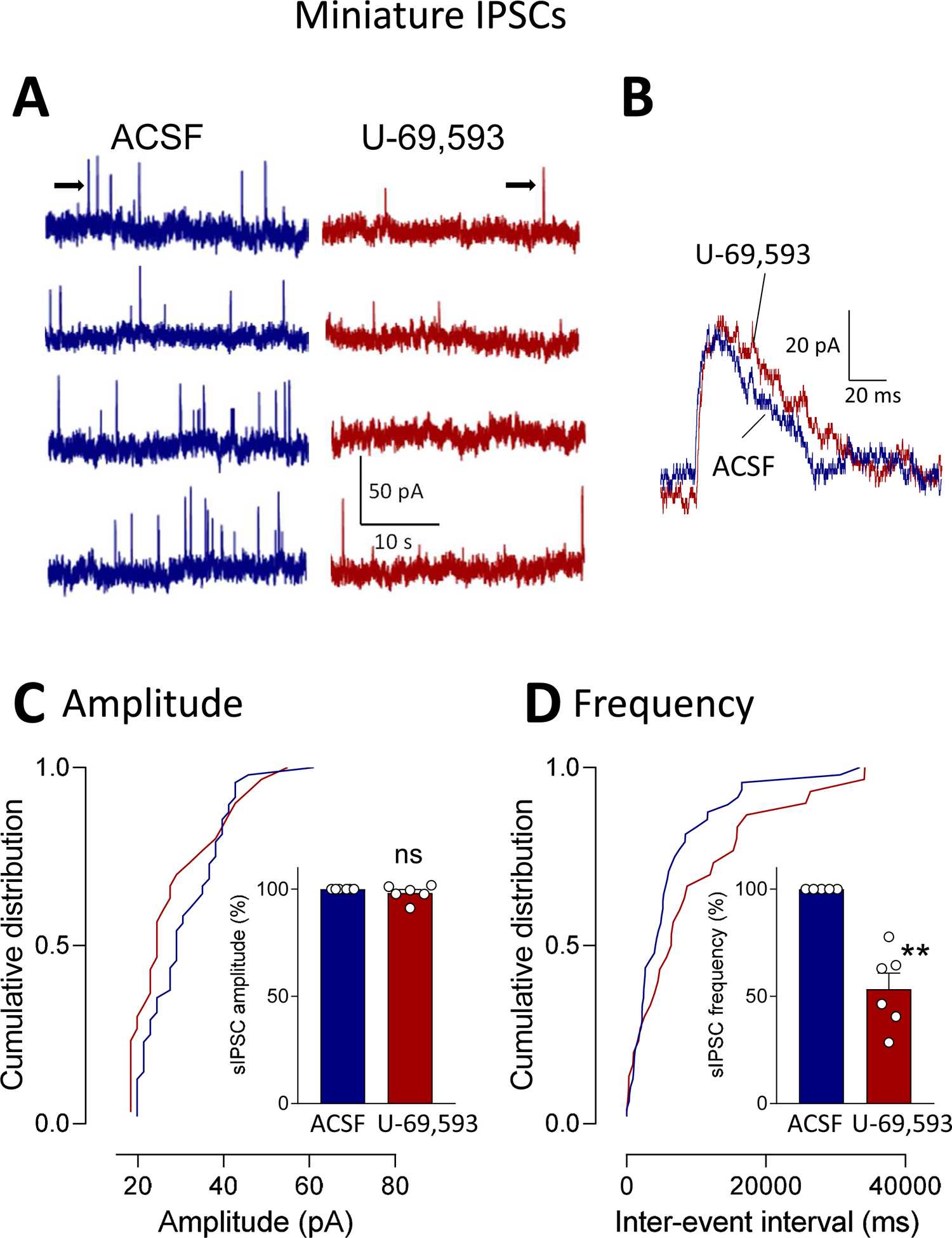
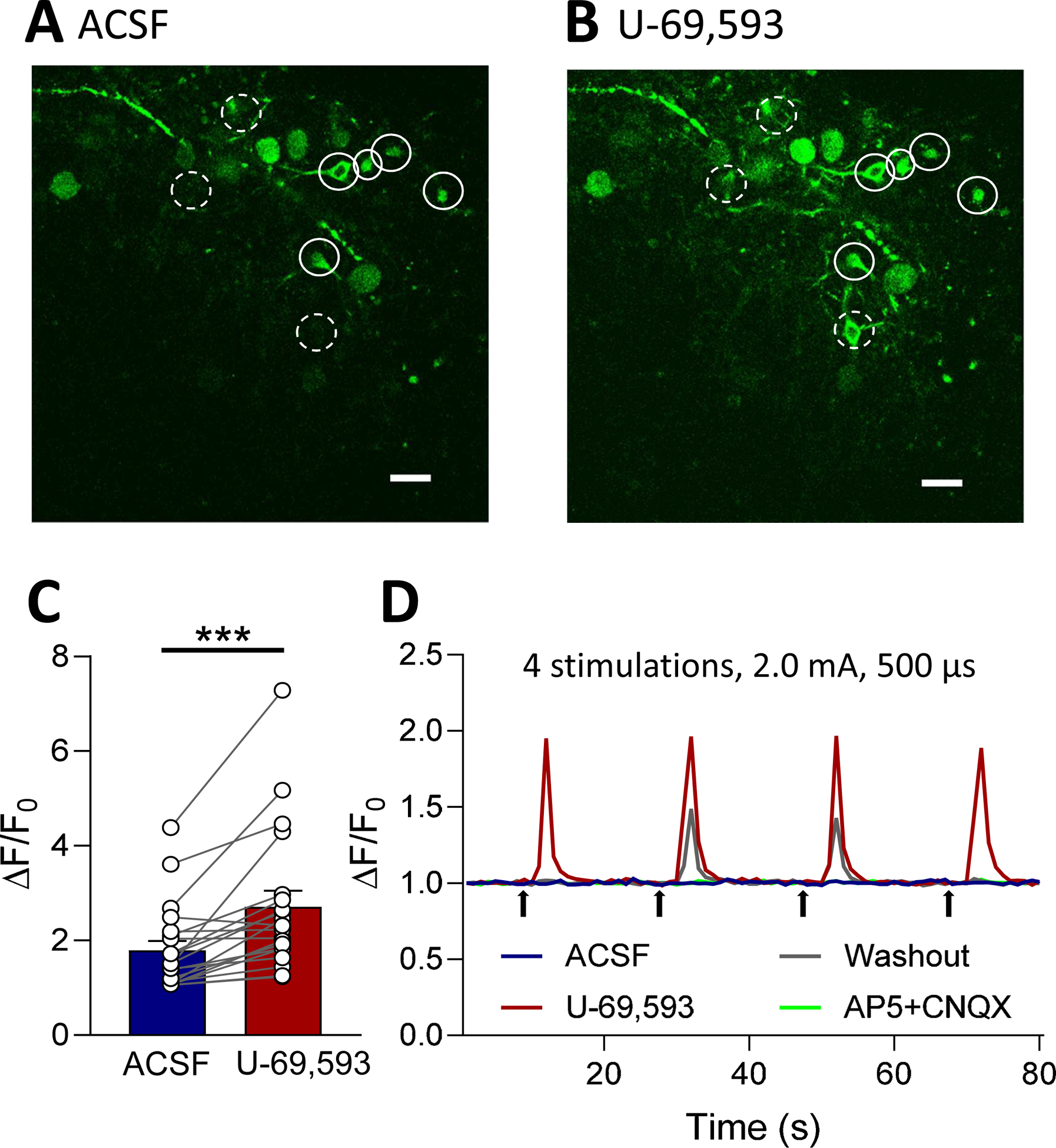
Recent evidence suggests that kappa opioid receptors (KOR) in limbic brain regions such as the amygdala contribute to pain conditions, but underlying mechanisms remain to be determined. The amygdala is an important player in averse-affective aspects of pain and pain modulation. The central nucleus (CeA) serves output functions through projection neurons that include corticotropin releasing factor (CRF) expressing neurons. The CeA is also rich in KOR. Here we tested the novel hypothesis that KOR activation in the CeA generates pain-like behaviors through a mechanism that involves inhibition of synaptic inhibition (disinhibition) of CRF neurons. Intra-CeA administration of a KOR agonist (U-69,593) increased vocalizations of naïve rats to noxious stimuli, and induced anxiety-like behaviors in the open field test (OFT) and avoidance in the conditioned place preference test, without affecting mechanosensory thresholds. Optogenetic silencing of CeA-CRF neurons blocked the facilitatory effects of systemically applied U-69,593 in naïve rats. Patch-clamp recordings of CRF neurons in rat brain slices found that U-69,593 decreased feedforward inhibitory transmission evoked by optogenetic stimulation of parabrachial afferents, but had no effect on monosynaptic excitatory transmission. U-69,593 decreased frequency, but not amplitude, of inhibitory synaptic currents, suggesting a presynaptic action. Multiphoton imaging of CeA-CRF neurons in rat brain slices showed that U-69,593 increased calcium signals evoked by electrical stimulation of presumed parabrachial input. This study shows for the first time that KOR activation increases activity of amygdala CRF neurons through synaptic disinhibition, resulting in averse-affective pain-like behaviors. Blocking KOR receptors may therefore represent a novel therapeutic strategy.
Keywords: Amygdala; Behavior; Corticotropin-releasing factor; Kappa opioid receptor; Pain; Patch clamp.
Copyright © 2021 Elsevier Ltd. All rights reserved.
Figures
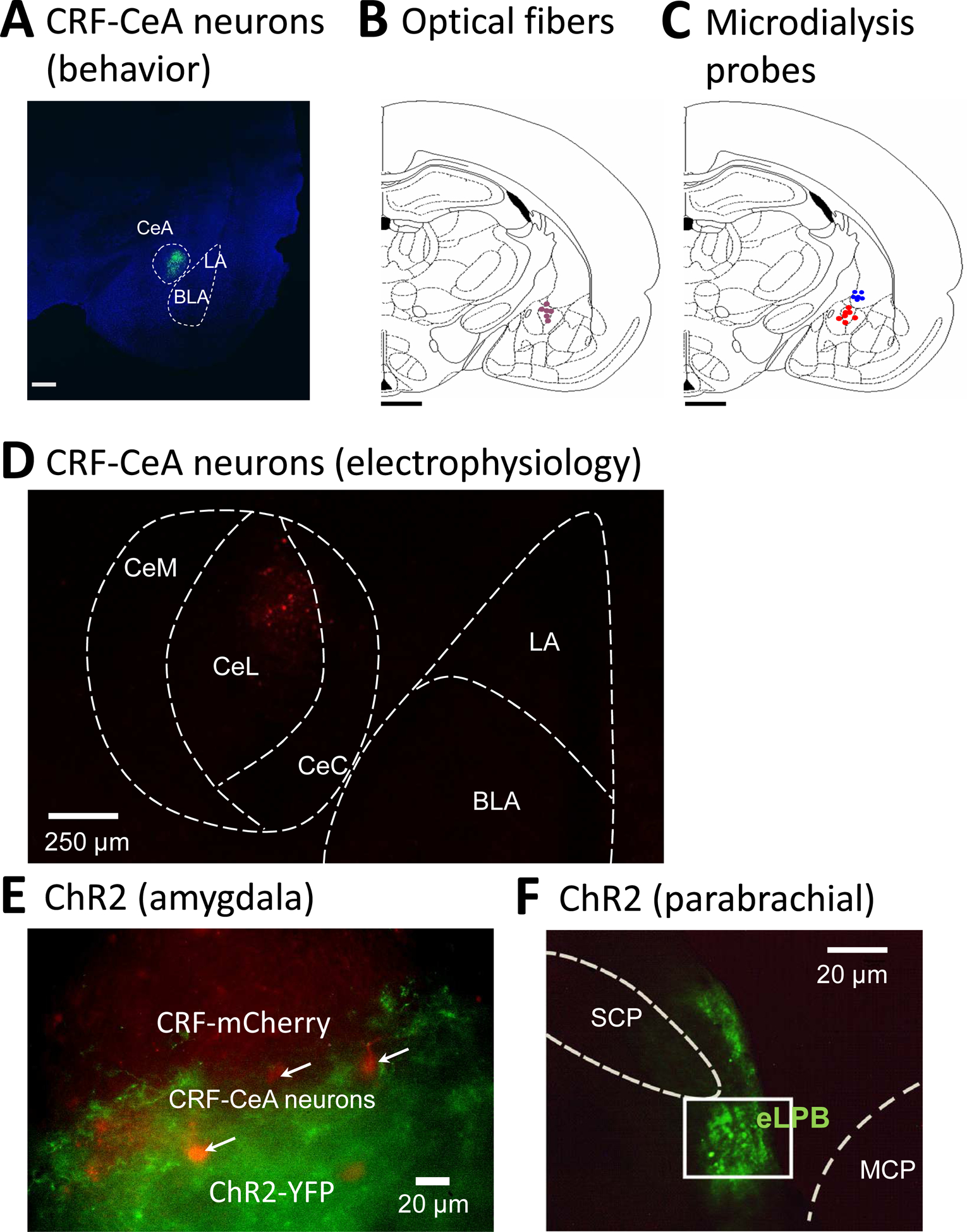
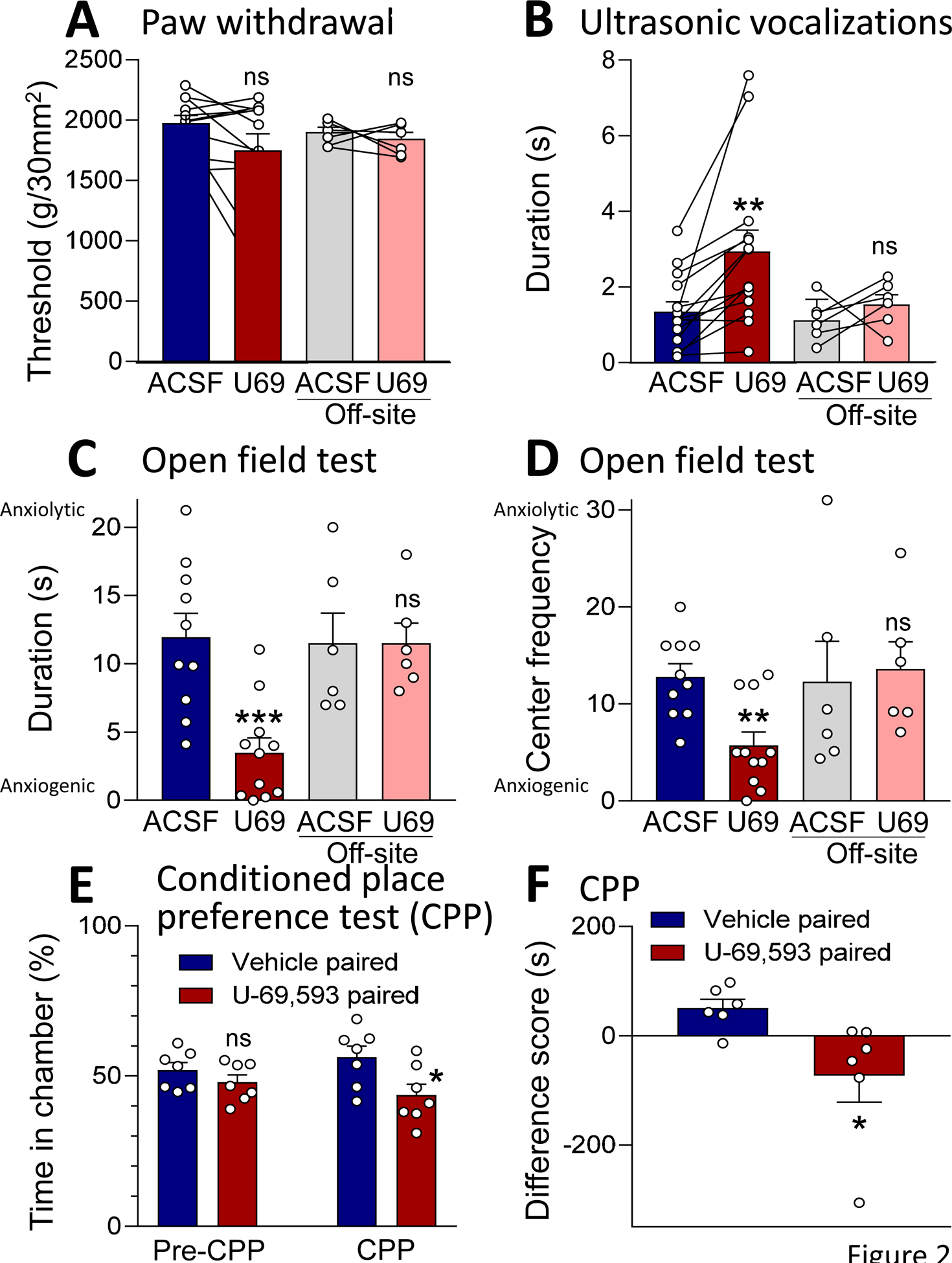
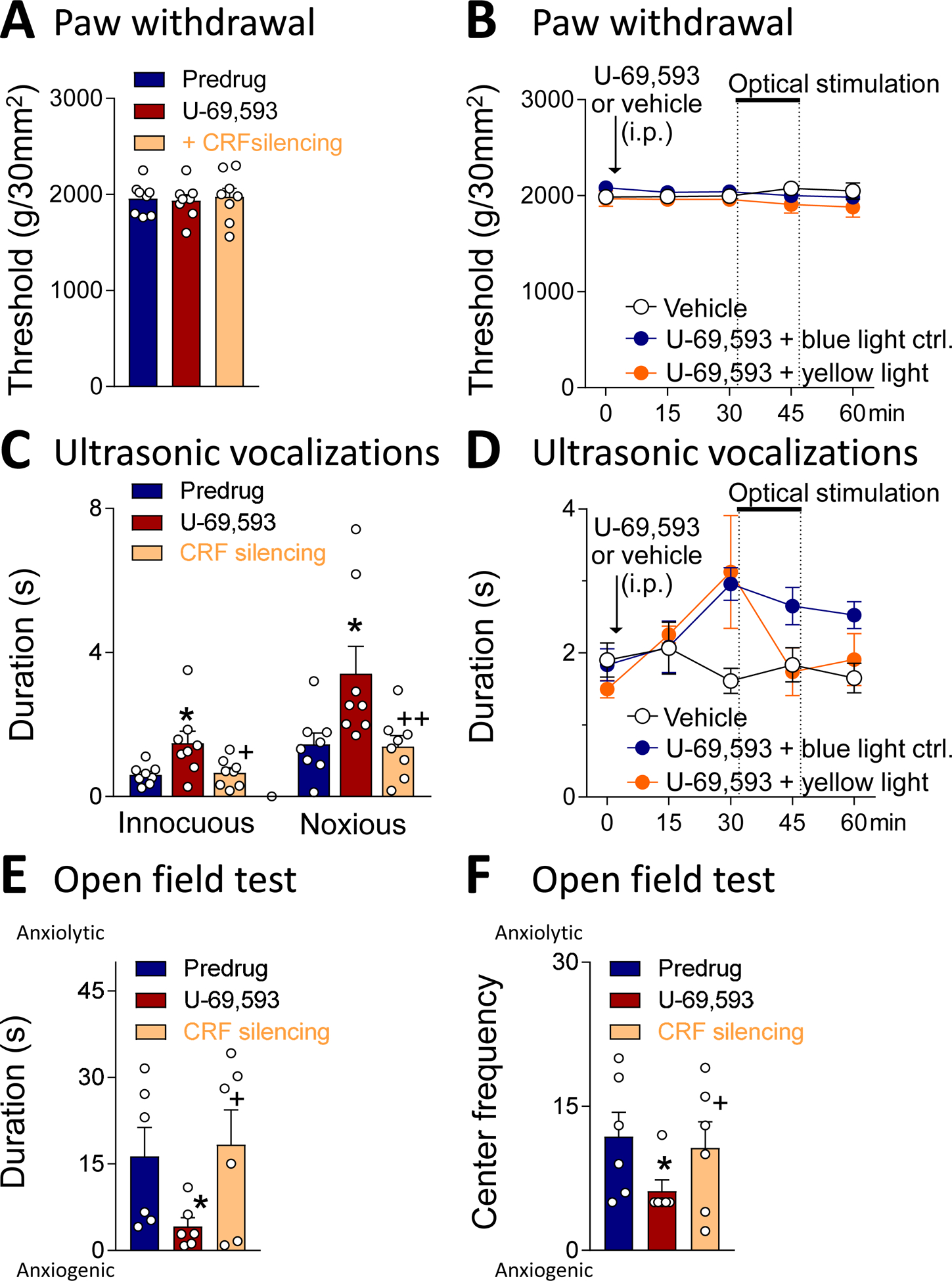
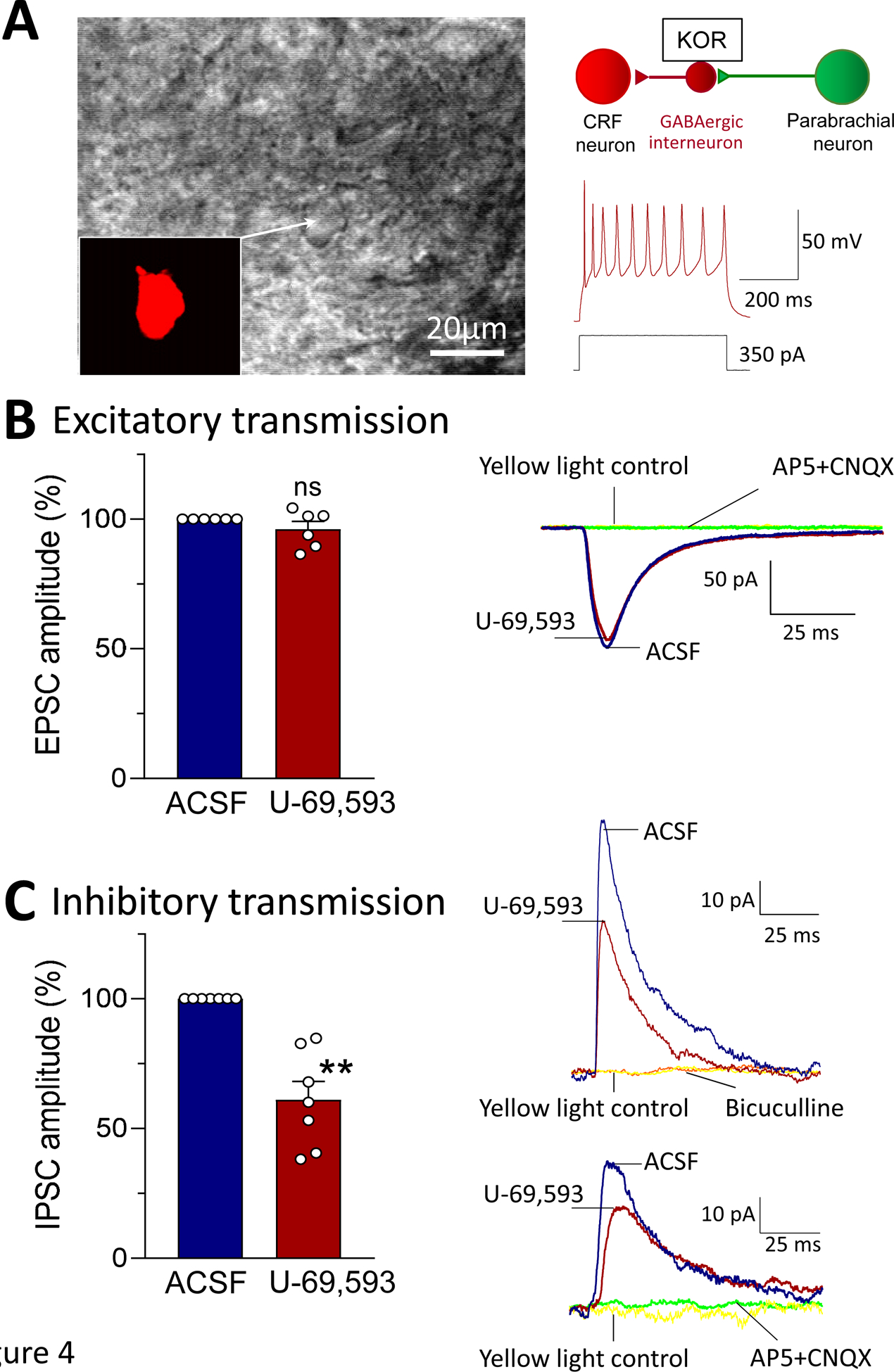
References
-
- Beckerman MA, Van Kempen TA, Justice NJ, Milner TA, Glass MJ, 2013. Corticotropin-releasing factor in the mouse central nucleus of the amygdala: ultrastructural distribution in NMDA-NR1 receptor subunit expressing neurons as well as projection neurons to the bed nucleus of the stria terminalis. Exp. Neurol 239, 120–132. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
