

Review of Venoms of Non-Polydnavirus Carrying Ichneumonoid Wasps
- PMID: 33445639
- PMCID: PMC7828074
- DOI: 10.3390/biology10010050
Review of Venoms of Non-Polydnavirus Carrying Ichneumonoid Wasps
Abstract
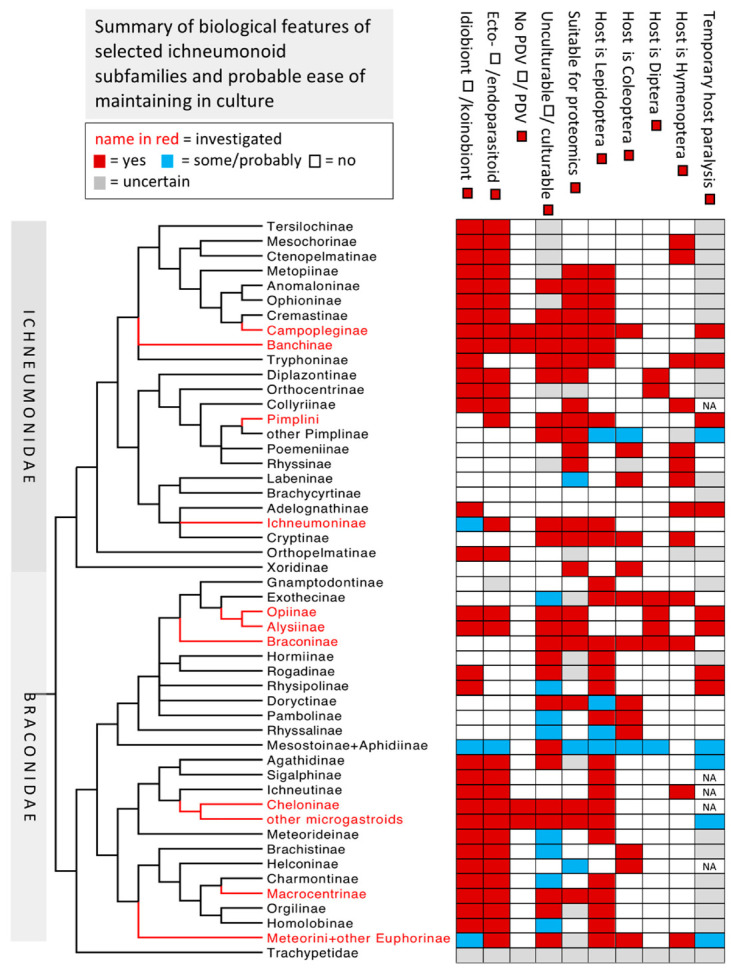
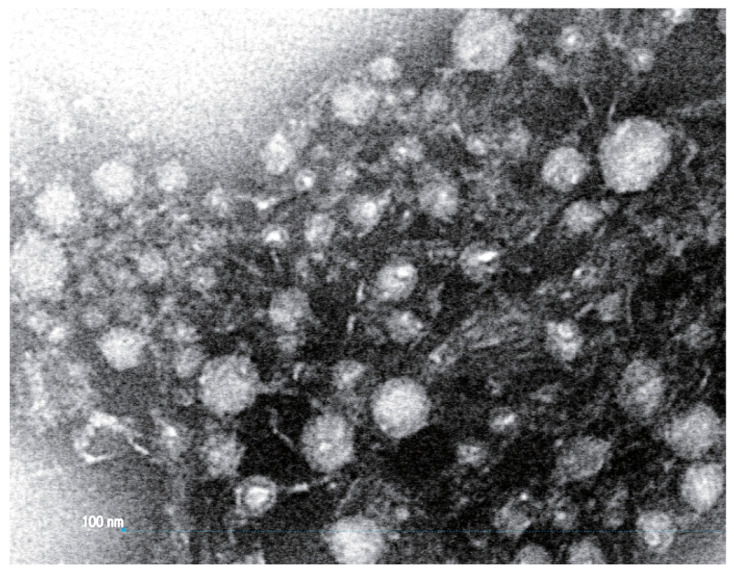
Parasitoids are predominantly insects that develop as larvae on or inside their host, also usually another insect, ultimately killing it after various periods of parasitism when both parasitoid larva and host are alive. The very large wasp superfamily Ichneumonoidea is composed of parasitoids of other insects and comprises a minimum of 100,000 species. The superfamily is dominated by two similarly sized families, Braconidae and Ichneumonidae, which are collectively divided into approximately 80 subfamilies. Of these, six have been shown to release DNA-containing virus-like particles, encoded within the wasp genome, classified in the virus family Polydnaviridae. Polydnaviruses infect and have profound effects on host physiology in conjunction with various venom and ovarial secretions, and have attracted an immense amount of research interest. Physiological interactions between the remaining ichneumonoids and their hosts result from adult venom gland secretions and in some cases, ovarian or larval secretions. Here we review the literature on the relatively few studies on the effects and chemistry of these ichneumonoid venoms and make suggestions for interesting future research areas. In particular, we highlight relatively or potentially easily culturable systems with features largely lacking in currently studied systems and whose study may lead to new insights into the roles of venom chemistry in host-parasitoid relationships as well as their evolution.
Keywords: Aphidius; Asobara; Braconidae; Habrobracon; Ichneumonidae; Pimpla.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
















Similar articles
-
[Co-evolutionary strategies of interaction between parasitoids and polydnaviruses].Rev Latinoam Microbiol. 2006 Jan-Mar;48(1):31-43. Rev Latinoam Microbiol. 2006. PMID: 17357572 Review. Spanish.
-
Phylogenomics of Ichneumonoidea (Hymenoptera) and implications for evolution of mode of parasitism and viral endogenization.Mol Phylogenet Evol. 2021 Mar;156:107023. doi: 10.1016/j.ympev.2020.107023. Epub 2020 Nov 28. Mol Phylogenet Evol. 2021. PMID: 33253830
-
Venom gland extract is not required for successful parasitism in the polydnavirus-associated endoparasitoid Hyposoter didymator (Hym. Ichneumonidae) despite the presence of numerous novel and conserved venom proteins.Insect Biochem Mol Biol. 2013 Mar;43(3):292-307. doi: 10.1016/j.ibmb.2012.12.010. Epub 2013 Jan 5. Insect Biochem Mol Biol. 2013. PMID: 23298679
-
Systematic analysis of a wasp parasitism arsenal.Mol Ecol. 2014 Feb;23(4):890-901. doi: 10.1111/mec.12648. Mol Ecol. 2014. PMID: 24383716 Free PMC article.
-
Modulation of immune responses to parasitoids by polydnaviruses.Parasitology. 1998;116 Suppl:S57-64. doi: 10.1017/s0031182000084948. Parasitology. 1998. PMID: 9695110 Review.
Cited by
-
Multi-omic approach to characterize the venom of the parasitic wasp Cotesia congregata (Hymenoptera: Braconidae).BMC Genomics. 2025 Apr 30;26(1):431. doi: 10.1186/s12864-025-11604-y. BMC Genomics. 2025. PMID: 40307720 Free PMC article.
-
Master of Puppets: How Microbiota Drive the Nematoda Ecology and Evolution?Ecol Evol. 2025 Aug 19;15(8):e71549. doi: 10.1002/ece3.71549. eCollection 2025 Aug. Ecol Evol. 2025. PMID: 40837528 Free PMC article. Review.
-
Venomous Cargo: Diverse Toxin-Related Proteins Are Associated with Extracellular Vesicles in Parasitoid Wasp Venom.Pathogens. 2025 Mar 5;14(3):255. doi: 10.3390/pathogens14030255. Pathogens. 2025. PMID: 40137740 Free PMC article.
-
Multiparasitism enables a specialist endoparasitoid to complete parasitism in an unsuitable host caterpillar.Sci Rep. 2025 Mar 11;15(1):8361. doi: 10.1038/s41598-025-91403-3. Sci Rep. 2025. PMID: 40069222 Free PMC article.
-
Effects of Parasitism and Venom from the Endoparasitoid Brachymeria lasus on Immunity of the Host Galleria mellonella.Insects. 2025 Aug 19;16(8):863. doi: 10.3390/insects16080863. Insects. 2025. PMID: 40870664 Free PMC article.
References
-
- Godfray H.C.J. Parasitoids: Behavioral and Evolutionary Ecology. Princeton University Press; Princeton, NJ, USA: 1994.
-
- Quicke D.L.J. Parasitic Wasps. Chapman & Hall; London, UK: 1997. pp. 1–470.
-
- Yu D.S.K., van Achterberg C., Horstmann K. Taxapad 2016, Ichneumonoidea 2015. [(accessed on 23 September 2017)];2016 Database on Flash-Drive, Nepean, ON, USA. Available online: http://www.taxapad.com.
-
- Quicke D.L.J., Austin A.D., Fagan-Jeffries E.P., Hebert P.D., Butcher B.A. Recognition of the Trachypetidae stat. n. as a new extant family of Ichneumonoidea (Hymenoptera), based on molecular and morphological evidence. Syst. Entomol. 2020;45:771–782.
-
- Quicke D.L.J. The Braconid and Ichneumonid Parasitic Wasps: Biology, Systematics, Evolution and Ecology. Wiley Blackwell; Oxford, UK: 2015. pp. 1–688.
Publication types
LinkOut - more resources
Full Text Sources
Other Literature Sources
