

Direct Observation of β-Barrel Intermediates in the Self-Assembly of Toxic SOD128-38 and Absence in Nontoxic Glycine Mutants
- PMID: 33445870
- PMCID: PMC7902411
- DOI: 10.1021/acs.jcim.0c01319
Direct Observation of β-Barrel Intermediates in the Self-Assembly of Toxic SOD128-38 and Absence in Nontoxic Glycine Mutants
Abstract
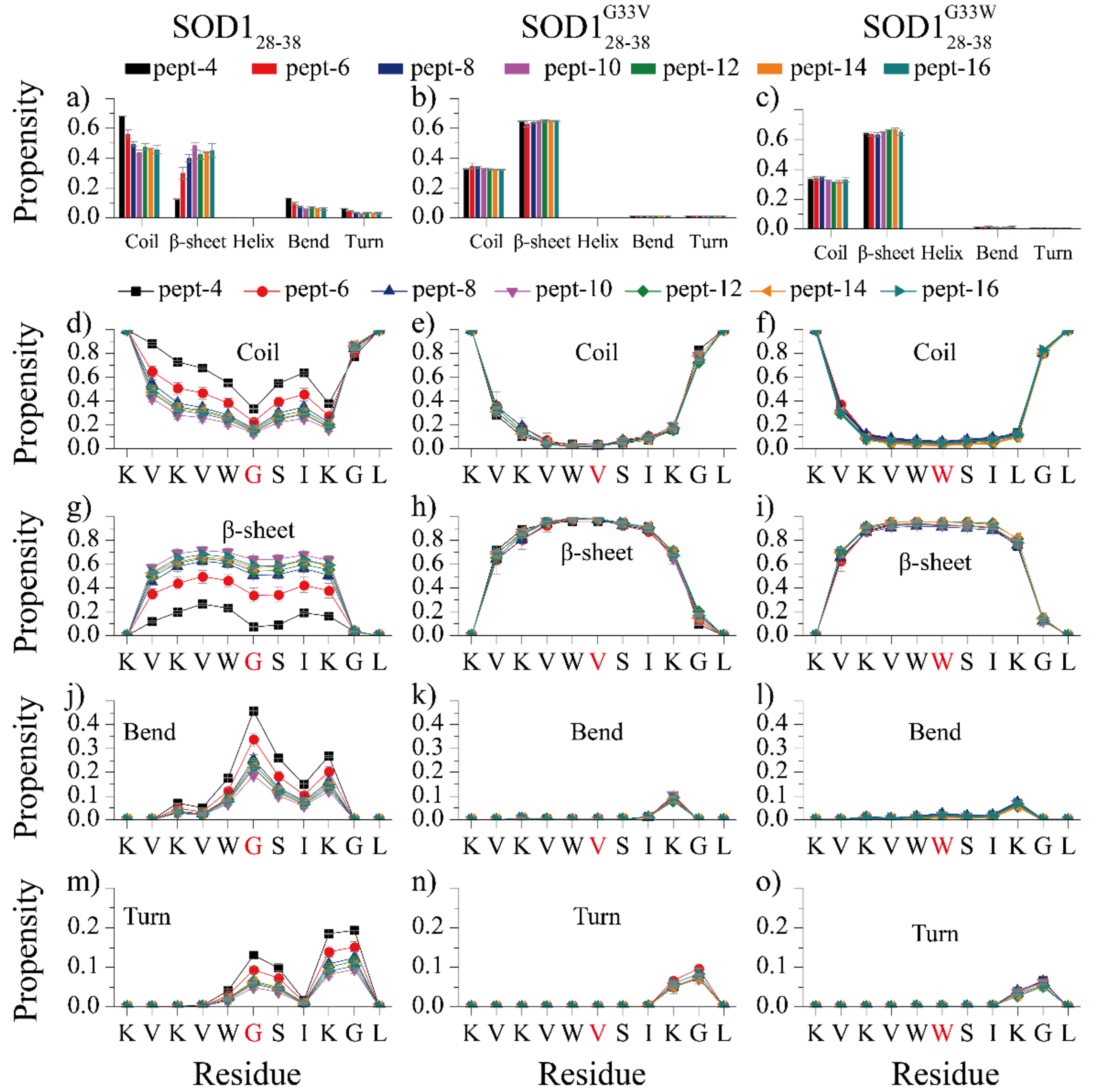
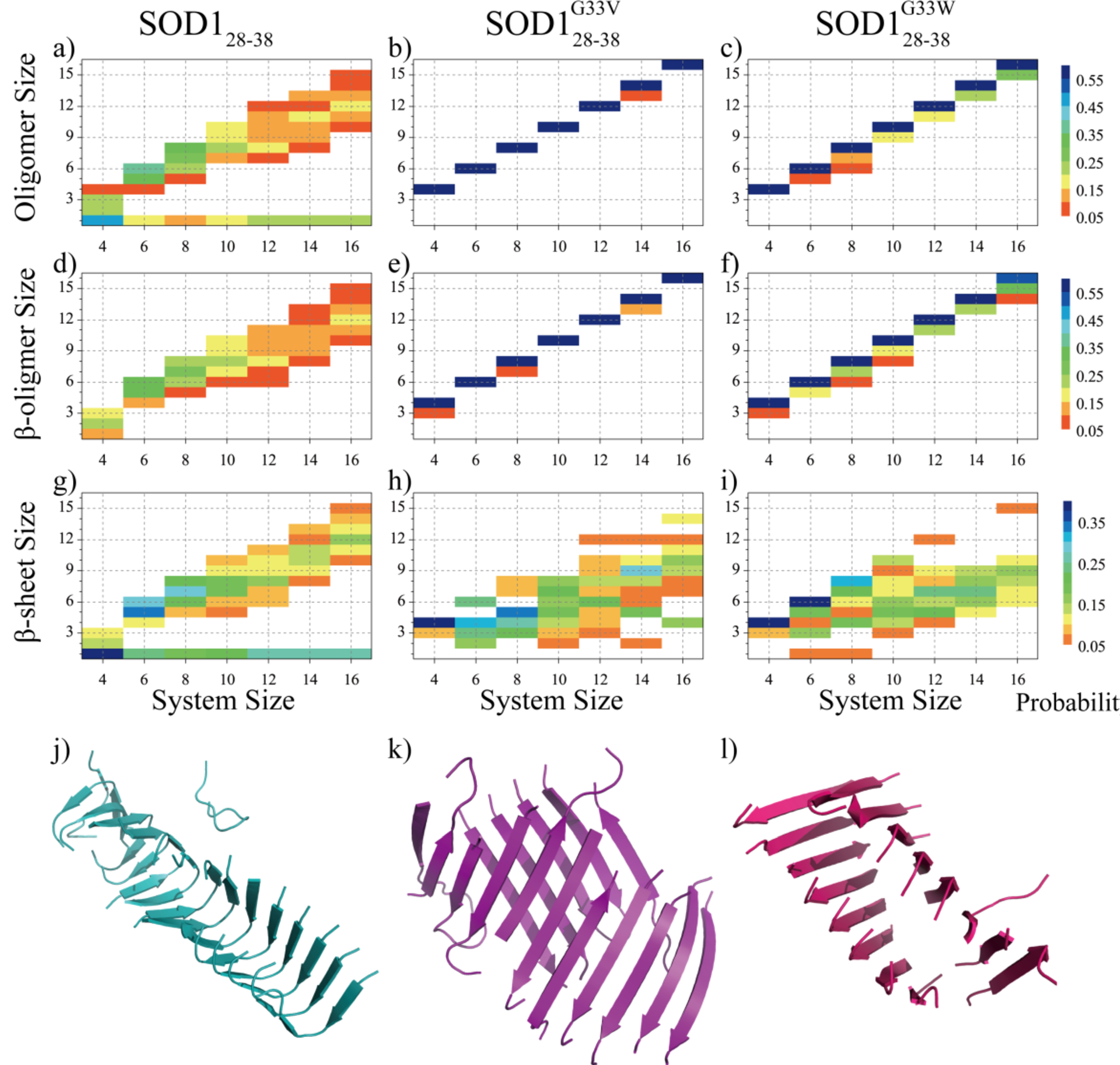
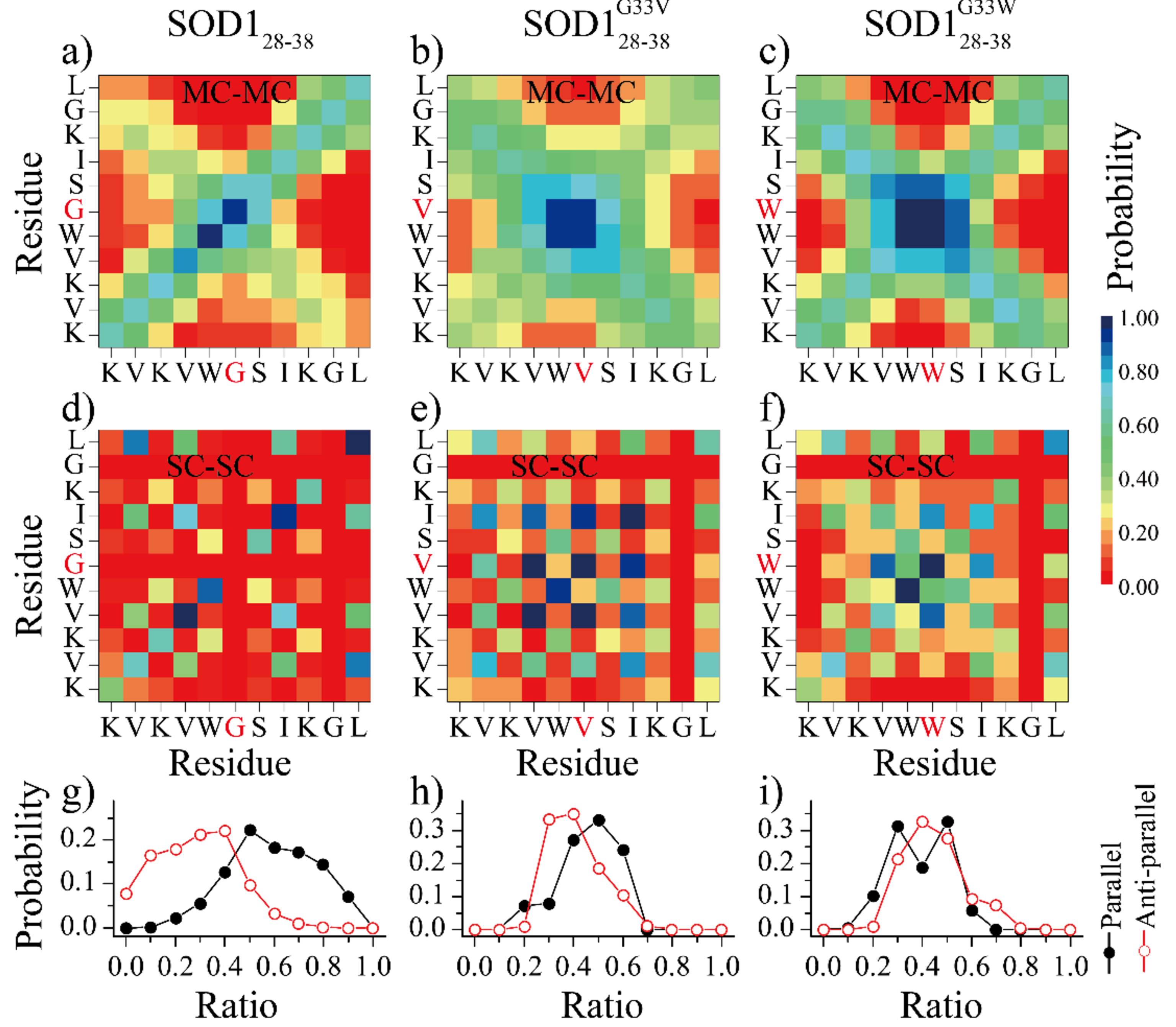
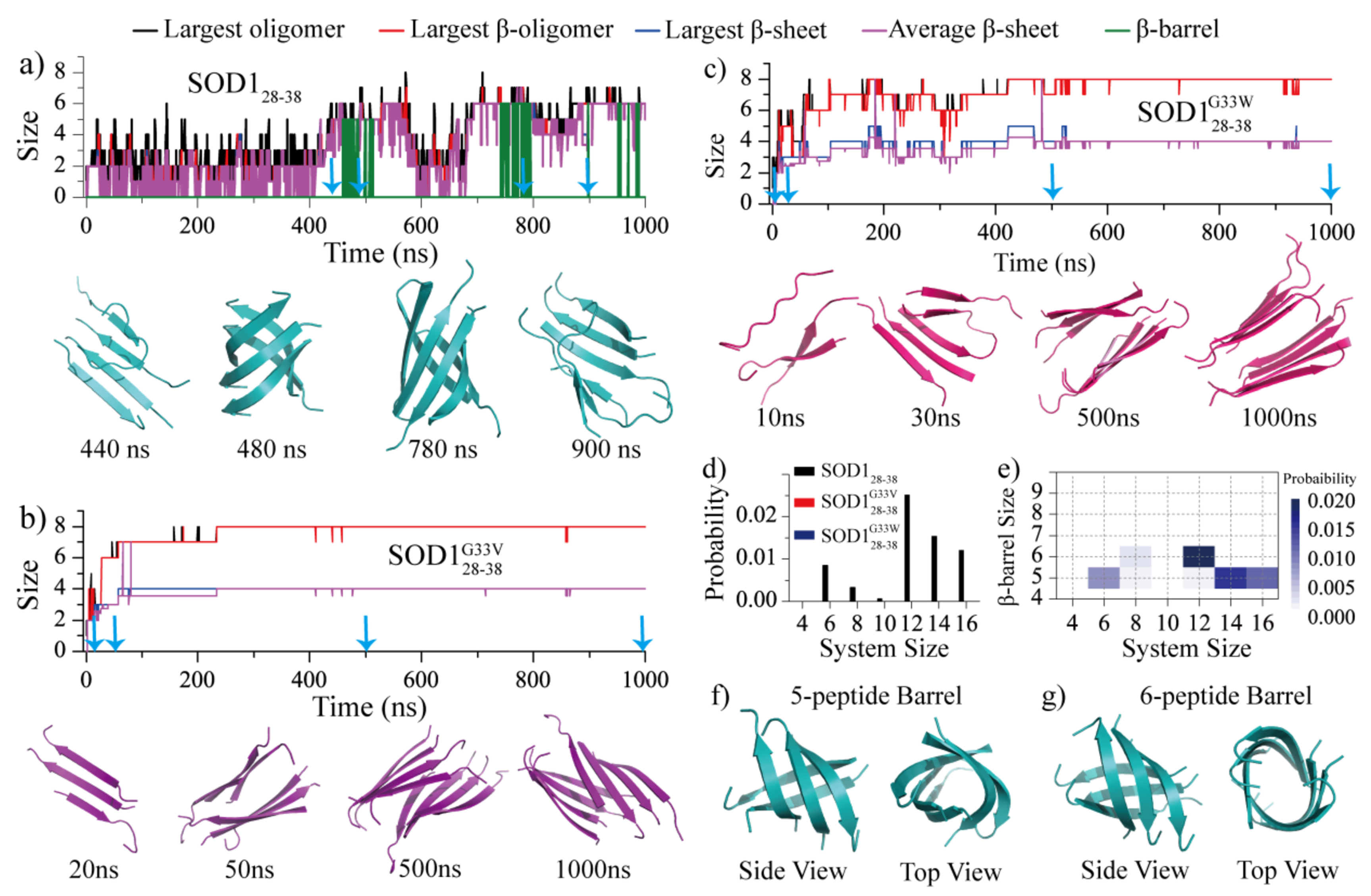
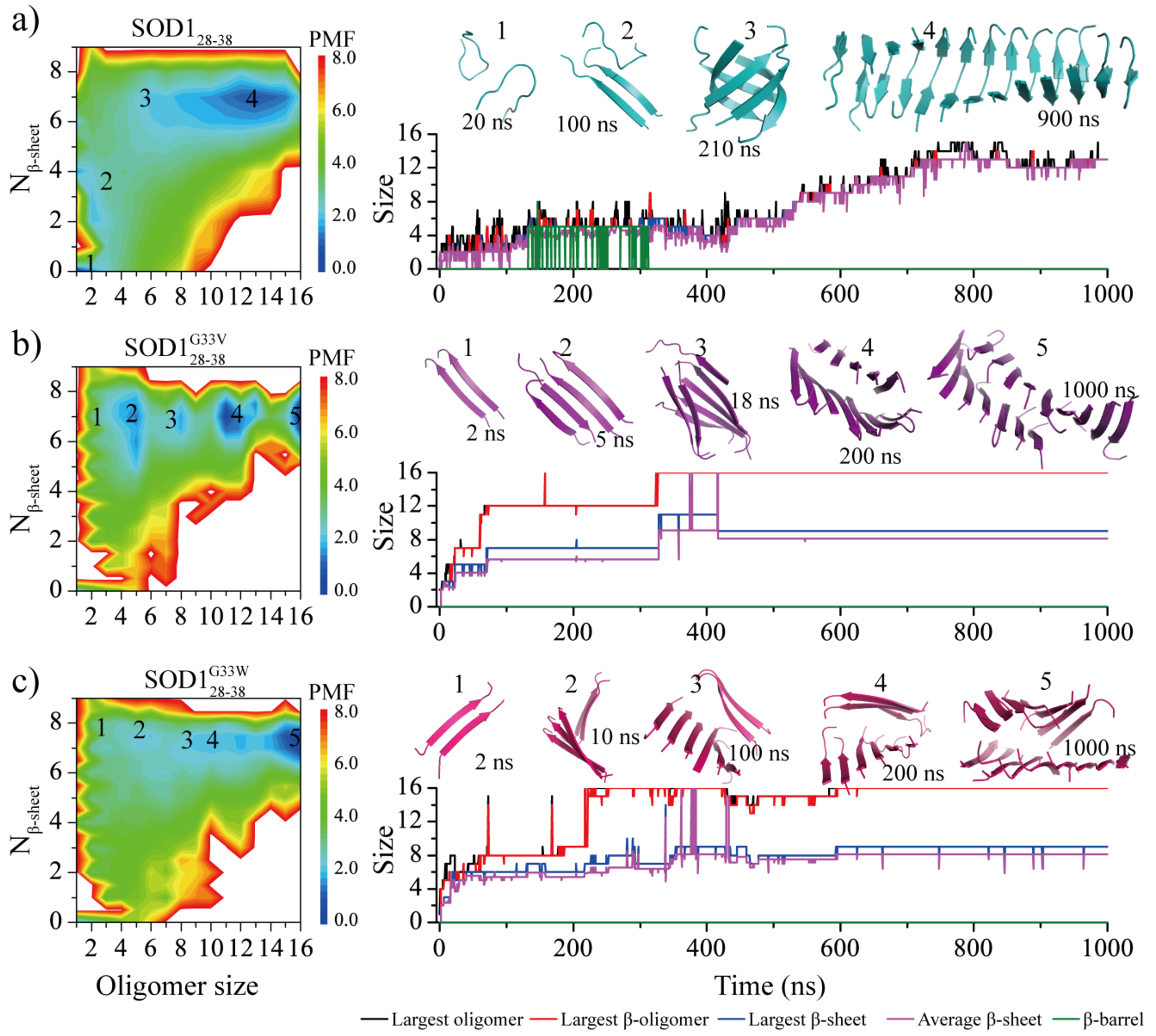
Soluble low-molecular-weight oligomers formed during the early stage of amyloid aggregation are considered the major toxic species in amyloidosis. The structure-function relationship between oligomeric assemblies and the cytotoxicity in amyloid diseases are still elusive due to the heterogeneous and transient nature of these aggregation intermediates. To uncover the structural characteristics of toxic oligomeric intermediates, we compared the self-assembly dynamics and structures of SOD128-38, a cytotoxic fragment of the superoxide dismutase 1 (SOD1) associated with the amyotrophic lateral sclerosis, with its two nontoxic mutants G33V and G33W using molecular dynamics simulations. Single-point glycine substitutions in SOD128-38 have been reported to abolish the amyloid toxicity. Our simulation results showed that the toxic SOD128-38 and its nontoxic mutants followed different aggregation pathways featuring distinct aggregation intermediates. Specifically, wild-type SOD128-38 initially self-assembled into random-coil-rich oligomers, among which fibrillar aggregates composed of well-defined curved single-layer β-sheets were nucleated via coil-to-sheet conversions and the formation of β-barrels as intermediates. In contrast, the nontoxic G33V/G33W mutants readily assembled into small β-sheet-rich oligomers and then coagulated with each other into cross-β fibrils formed by two-layer β-sheets without forming β-barrels as the intermediates. The direct observation of β-barrel oligomers during the assembly of toxic SOD128-38 fragments but not the nontoxic glycine-substitution mutants strongly supports β-barrels as the toxic oligomers in amyloidosis, probably via interactions with the cell membrane and forming amyloid pores. With well-defined structures, the β-barrel might serve as a novel therapeutic target against amyloid-related diseases.
Conflict of interest statement
Conflicts of interest
There are no conflicts to declare.
Figures
Similar articles
-
β-barrel Oligomers as Common Intermediates of Peptides Self-Assembling into Cross-β Aggregates.Sci Rep. 2018 Jul 9;8(1):10353. doi: 10.1038/s41598-018-28649-7. Sci Rep. 2018. PMID: 29985420 Free PMC article.
-
Nucleation of β-rich oligomers and β-barrels in the early aggregation of human islet amyloid polypeptide.Biochim Biophys Acta Mol Basis Dis. 2019 Feb 1;1865(2):434-444. doi: 10.1016/j.bbadis.2018.11.021. Epub 2018 Nov 28. Biochim Biophys Acta Mol Basis Dis. 2019. PMID: 30502402 Free PMC article.
-
Structures and dynamics of β-barrel oligomer intermediates of amyloid-beta16-22 aggregation.Biochim Biophys Acta Biomembr. 2018 Sep;1860(9):1687-1697. doi: 10.1016/j.bbamem.2018.03.011. Epub 2018 Mar 14. Biochim Biophys Acta Biomembr. 2018. PMID: 29550287 Free PMC article.
-
β-Barrels and Amyloids: Structural Transitions, Biological Functions, and Pathogenesis.Int J Mol Sci. 2021 Oct 20;22(21):11316. doi: 10.3390/ijms222111316. Int J Mol Sci. 2021. PMID: 34768745 Free PMC article. Review.
-
Elucidating the Structures of Amyloid Oligomers with Macrocyclic β-Hairpin Peptides: Insights into Alzheimer's Disease and Other Amyloid Diseases.Acc Chem Res. 2018 Mar 20;51(3):706-718. doi: 10.1021/acs.accounts.7b00554. Epub 2018 Mar 6. Acc Chem Res. 2018. PMID: 29508987 Free PMC article. Review.
Cited by
-
Structural Perturbation of Monomers Determines the Amyloid Aggregation Propensity of Calcitonin Variants.J Chem Inf Model. 2023 Jan 9;63(1):308-320. doi: 10.1021/acs.jcim.2c01202. Epub 2022 Dec 1. J Chem Inf Model. 2023. PMID: 36456917 Free PMC article.
-
Molecular Insights into the Self-Assembly of Block Copolymer Suckerin Polypeptides into Nanoconfined β-Sheets.Small. 2022 Aug;18(34):e2202642. doi: 10.1002/smll.202202642. Epub 2022 Jul 28. Small. 2022. PMID: 35901284 Free PMC article.
-
Computational Investigation of Coaggregation and Cross-Seeding between Aβ and hIAPP Underpinning the Cross-Talk in Alzheimer's Disease and Type 2 Diabetes.J Chem Inf Model. 2024 Jul 8;64(13):5303-5316. doi: 10.1021/acs.jcim.4c00859. Epub 2024 Jun 26. J Chem Inf Model. 2024. PMID: 38921060 Free PMC article.
-
Misfolding and Self-Assembly Dynamics of Microtubule-Binding Repeats of the Alzheimer-Related Protein Tau.J Chem Inf Model. 2021 Jun 28;61(6):2916-2925. doi: 10.1021/acs.jcim.1c00217. Epub 2021 May 25. J Chem Inf Model. 2021. PMID: 34032430 Free PMC article.
-
Computational insights into the cross-talk between medin and Aβ: implications for age-related vascular risk factors in Alzheimer's disease.Brief Bioinform. 2024 Jan 22;25(2):bbad526. doi: 10.1093/bib/bbad526. Brief Bioinform. 2024. PMID: 38271485 Free PMC article.
References
-
- Iadanza MG; Jackson MP; Hewitt EW; Ranson NA; Radford SE, A new era for understanding amyloid structures and disease. Nature reviews. Molecular cell biology 2018, 19, 755–773. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
